Prokarüootne rakk on palju lihtsam kui loomade ja taimede rakud. Väljast on see kaetud rakuseinaga, mis täidab kaitse-, vormimis- ja transpordifunktsioone. Rakuseina jäikuse annab mureiin. Mõnikord on bakterirakk pealt kaetud kapsli või limakihiga.
Bakterite protoplasma, nagu eukarüootidelgi, on ümbritsetud plasmamembraan. Hingamisprotsessis osalevaid mesosoome, bakterioklorofülli ja muid pigmente leidub membraani kotitaolistes, torukujulistes või lamellsetes invaginatsioonides. Prokarüootide geneetiline materjal ei moodusta tuuma, vaid paikneb otse tsütoplasmas. Bakteriaalne DNA on üks ümmargune molekul, millest igaüks koosneb tuhandetest ja miljonitest aluspaaridest. Bakteriraku genoom on palju lihtsam kui arenenumate olendite rakkudel: keskmiselt sisaldab bakteri DNA mitu tuhat geeni.
Ei leitud prokarüootsetes rakkudes endoplasmaatiline retikulum, a ribosoomid hõljuvad vabalt tsütoplasmas. Mitte prokarüootides mitokondrid; osa nende funktsioonist täidab rakumembraan.
prokarüootid
Bakterid on rakulise struktuuriga organismidest väikseimad; nende suurus on vahemikus 0,1 kuni 10 µm. Tüüpiline trükipunkt mahutab sadu tuhandeid keskmise suurusega baktereid. Baktereid saab näha ainult läbi mikroskoobi, mistõttu neid nimetatakse mikroorganismid või mikroobid; uuritakse mikroorganisme mikrobioloogia . Mikrobioloogia osa, mis tegeleb bakteritega, nimetatakse bakterioloogia . Selle teaduse algus pandi Anthony van Leeuwenhoek 17. sajandil.
bakterid on vanimad teadaolevad organismid. Bakterite ja sinivetikate (stromatoliitide) elutegevuse jäljed kuuluvad Arheani ja ulatuvad 3,5 miljardi aasta taha.Erinevate liikide ja isegi sugukondade esindajate vahelise geenivahetuse võimaluse tõttu on prokarüoote üsna keeruline süstematiseerida. Prokarüootide rahuldav taksonoomia ei ole veel üles ehitatud; kõik olemasolevad süsteemid on kunstlikud ja klassifitseerivad baktereid mõne märgirühma järgi, arvestamata nende fülogeneetilist seost. Varem bakterid koos seened ja vetikad kuuluvad madalamate taimede alamkuningriiki. Bakterid liigitatakse praegu eraldi prokarüootide kuningriigiks. Kõige tavalisem klassifikatsioonisüsteem on Bergi süsteem põhineb rakuseina struktuuril.
20. sajandi lõpus avastasid teadlased, et suhteliselt vähe uuritud bakterirühma rakud - arhebakterid - sisaldama rRNA, mis erinevad oma struktuuri poolest nii prokarüootsest rRNA-st kui ka eukarüootsest rRNA-st. Arhebakterite geneetilise aparaadi struktuur (esinemine intronid ja korduvad järjestused töötlemine, vorm ribosoom) lähendab neid eukarüootidele; teisalt on arhebakteritel ka tüüpilised prokarüootidele iseloomulikud tunnused (tuuma puudumine rakus, flagellade, plasmiidide ja gaasivakuoolide olemasolu, rRNA suurus, lämmastiku sidumine). Lõpuks erinevad arhebakterid kõigist teistest organismidest rakuseina struktuuri, fotosünteesi tüübi ja mõnede muude omaduste poolest. Arhebakterid on võimelised eksisteerima ekstreemsetes tingimustes (näiteks kuumaveeallikates temperatuuril üle 100 ° C, ookeani sügavustes rõhul 260 atm, küllastunud soolalahustes (30% NaCl)). Mõned arhebakterid toodavad metaani, teised aga kasutavad energia saamiseks väävliühendeid.
Ilmselt on arhebakterid väga iidne organismide rühm; "äärmuslikud" võimalused annavad tunnistust Maa pinnale iseloomulikest tingimustest aastal arhea ajastu. Arvatakse, et arhebakterid on kõige lähemal hüpoteetilistele "rakkudele", mis hiljem põhjustasid kogu elu mitmekesisuse Maal.
Viimasel ajal on selgunud, et on kolm peamist tüüpi rRNA Esitatakse vastavalt esimene - eukarüootsetes rakkudes, teine - tõeliste bakterite rakkudes, samuti mitokondrid ja kloroplastid eukarüootides, kolmas - arhebakterites. Molekulaargeneetika uuringud on sundinud eukarüootide päritolu teooriat uue pilguga vaatama. Praegu arvatakse, et iidsel Maal arenesid üheaegselt kolm erinevat prokarüootide haru – arhebakterid, eubakterid ja urkarüootid , mida iseloomustab erinev struktuur ja erinevad energia saamise viisid. Urkarüootid, mis olid tegelikult eukarüootide tuuma-tsütoplasmaatiline komponent, lisati hiljem kui sümbionte erinevate eubakterite rühmade esindajad, mis muutusid tulevaste eukarüootsete rakkude mitokondriteks ja kloroplastideks.
Seega on arhebakterite jaoks varem määratud klassijärk selgelt ebapiisav. Praegu kipuvad paljud teadlased jagama prokarüoote kahte kuningriiki: arhebakterid ja tõelised bakterid (eubakterid ) või isegi eraldada arhebakterid eraldi Arhea kuningriiki.
Tõeliste bakterite klassifikatsioon on toodud skeem.
AT bakterirakk tuum puudub, kromosoomid paiknevad vabalt tsütoplasmas. Lisaks puuduvad bakterirakus membraani organellid: mitokondrid, EPS, golgi aparaat jne Väljaspool on rakumembraan kaetud rakuseinaga.
Enamik baktereid liigub passiivselt, kasutades vee- või õhuvoolu. Ainult mõnel neist on liikumisorganellid - flagella . Prokarüootsed lipukesed on ehituselt väga lihtsad ja koosnevad flagelliinvalgust, mis moodustab 10–20 nm läbimõõduga õõnsa silindri. Nad kruvivad söötmesse, liigutades rakku edasi. Ilmselt on see ainuke looduses teadaolev struktuur, mis kasutab ratta põhimõtet.
Bakterid jagunevad nende kuju järgi mitmeks rühmaks:
cocci (on ümar kuju);
batsillid (on vardakujuline vorm);
spirilla (on spiraali kujuga);
vibrios (olema koma kujul).
Hingamisviisi järgi jagunevad bakterid aeroobid (enamik baktereid) ja anaeroobid (teetanuse, botulismi, gaasigangreeni tekitajad). Esimesed vajavad hingamiseks hapnikku, teise jaoks on hapnik kasutu või isegi mürgine.
Bakterid paljunevad jagunedes umbes iga 20 minuti järel (soodsates tingimustes). DNA replitseerub, iga tütarrakk saab oma algse DNA koopia. Samuti on võimalik DNA-d üle kanda mittejagunevate rakkude vahel (läbi "palja" DNA püüdmise, abiga bakteriofaagid või poolt konjugatsioonid , kui bakterid on omavahel seotud kopuleerivate fimbriatega), siis isendite arvu suurenemist ei toimu. Paljunemist takistavad päikesekiired ja nende enda elutegevuse saadused.Bakterite käitumine pole eriti keeruline. Keemilised retseptorid registreerivad muutusi keskkonna happesuses ja erinevate ainete kontsentratsioonis: suhkrud, aminohapped, hapnik. Paljud bakterid reageerivad temperatuuri või valguse muutustele ning mõned bakterid võivad tajuda Maa magnetvälja.
Ebasoodsates tingimustes on bakter kaetud tiheda kestaga, tsütoplasma on dehüdreeritud ja elutegevus peaaegu peatub. Selles olekus võivad bakterite eosed viibida tunde sügavas vaakumis, taluda temperatuure vahemikus -240 ° C kuni +100 ° C.
BAKTERITE ÜLDOMADUSED
Rakulise struktuuriga organismid jagunevad kahte rühma: eukarüootid ja prokarüootid.
eukarüootid(kreeka keelest. eu- hea ja karyon- tuum) - organismid, mis sisaldavad rakkudes selgelt määratletud tuuma. Eukarüootide hulka kuuluvad ühe- ja mitmerakulised taimed, seened ja loomad, st kõik organismid, välja arvatud bakterid. Erinevate kuningriikide eukarüootsed rakud erinevad mitmel viisil. Kuid paljudes aspektides on nende struktuur sarnane. Millised on eukarüootsete rakkude omadused?
Loomarakkudel puudub rakumembraan, mis on taimedel ja seentel, samuti puuduvad plastiidid, mis taimedel ja mõnel bakteril on. Loomarakkude vakuoolid on väga väikesed ja ebastabiilsed. Kõrgematel taimedel pole tsentriole leitud.
Rakud prokarüootid(alates lat. umbes asemel, ees ja karüoot) ei sisalda formaliseeritud tuuma. Nende tuumaaine asub tsütoplasmas ja ei ole sellest membraaniga piiritletud. Prokarüootid on kõige iidsemad primitiivsed üherakulised organismid. Nende hulka kuuluvad bakterid ja tsüanobakterid (joonis 1). Nad paljunevad lihtsa jagunemise teel. Prokarüootidel sisaldab tsütoplasma ühte ringikujulist DNA molekuli, mida nimetatakse nukleoid või bakteriaalne kromosoom. Ribosoomid asuvad otse tsütoplasmas. Prokarüootsed rakud on haploidsed. Need ei sisalda mitokondreid, Golgi kompleksi, EPS-i. ATP süntees toimub neis plasmamembraanil.
Eriline koht eluslooduses on hõivatud viirused. Neil puudub rakuline struktuur ja need koosnevad nukleiinhappemolekulist – DNA-st või RNA-st, mida ümbritsevad kestana valgumolekulid.
Viirused põhjustavad mitmeid taimede, seente, loomade ja inimeste haigusi. Näiteks tubaka mosaiikviirus siseneb tubakalehtede rakkudesse, hävitab klorofülli ja leht muutub laiguliseks. Tuntud on inimese viirushaigused: rõuged, gripp, leetrid, poliomüeliit, marutaudi jne.
Riis. 10. Bakterirakkude (A) ja tsüanobakterite (B) struktuuri skeem:
1 - rakumembraan, 2 - kromosoom, 3 - tsütoplasma, 4 - plasmamembraan, 5 - ribosoom, 6 - säilitusained, 7 - flagellad.
Riis. 11. Tubaka mosaiikviirus:
I - haigusest mõjutatud tubakaleht, II - viiruse kristall rakus, III - tubaka mosaiikviiruse struktuuri skeem;
1 - valgumolekulide kest, 2 - spiraaliks keritud RNA ahel.
Ülesanded ja testid teemal "Teema 3. "Prokarüootne rakk. Viirused."
- Raku keemiline koostis
Tunnid: 8 Ülesanded: 10 Viktoriinid: 1
- taimerakk - Taimede rakuline struktuur Bakterid. Seened. Taimed (5.–6. klass)
Tunnid: 1 Ülesanded: 7 Kontrolltööd: 1
- Elusorganismide rakkude ehituse sarnasused ja erinevused - Tsütoloogia – rakkude teadus Üldised bioloogilised mustrid (9.–11. klass)
Tunnid: 2 Ülesanded: 11 Kontrolltööd: 1
- Rakuteooria. Rakuorganellid, nende funktsioonid - Tsütoloogia – rakkude teadus Üldised bioloogilised mustrid (9.–11. klass)
- Nimetage struktuuride funktsioonid ja märkige, millistes rakkudes (taime-, looma- või prokarüootses) need asuvad: tuum, tuumamembraan, nukleoplasma, kromosoomid, plasmamembraan, ribosoom, mitokondrid, rakusein, kloroplast, vakuool, lüsosoom, sile endoplasmaatiline retikulum ( agranulaarne) ja töötlemata (granulaarne), rakukeskus, golgi aparaat, tsilium, flagellum, mesosoom, pili või fimbriae.
- Nimeta vähemalt kolm märki, mille järgi saab taimerakku loomarakust eristada.
- Loetlege peamised erinevused prokarüootsete ja eukarüootsete rakkude vahel.
- Teema 1. "Plasmamembraan." §1, §8 lk 5;20
- Teema 2. "Puur". §8-10 lk 20-30
- Teema 3. "Prokarüootne rakk. Viirused." §11 lk 31-34
Ivanova T.V., Kalinova G.S., Myagkova A.N. "Üldbioloogia". Moskva, "Valgustus", 2000
prokarüootsed rakud- need on kõige primitiivsemad, väga lihtsalt korraldatud, säilitades iidsete aegade tunnused. To prokarüootne(või tuumaeelsete) organismide hulka kuuluvad bakterid ja sinivetikad (tsüanobakterid). Struktuuri ühtsuse ja teravate erinevuste alusel teistest rakkudest eraldatakse prokarüootsed rakud iseseisvaks jahipüsside kuningriigiks.
Mõelge struktuurile prokarüootne rakk bakterite näitel. Prokarüootse raku geneetilist aparaati esindab ühe tsükli kromosoomi DNA, mis asub tsütoplasmas ja ei ole sellest membraaniga piiritletud. Sellist tuuma analoogi nimetatakse nukleoidiks. DNA ei moodusta valkudega komplekse ja seetõttu "töötavad" kõik kromosoomi moodustavad geenid, s.t. neist loetakse pidevalt teavet.
prokarüootne rakkümbritsetud membraaniga, mis eraldab tsütoplasmat rakuseinast ja on moodustunud keerulisest kõrge polümeerisisaldusega ainest. Tsütoplasmas on vähe organelle, kuid leidub arvukalt väikseid ribosoome (bakterirakud sisaldavad 5000–50 000 ribosoomi).
Prokarüootse raku struktuur
Prokarüootse raku tsütoplasma on läbi imbunud membraanidega, mis moodustavad endoplasmaatilise retikulumi, ja see sisaldab ribosoome, mis teostavad valgusünteesi.
Prokarüootse raku rakuseina sisemist osa esindab plasmamembraan, mille eendid tsütoplasmasse moodustavad mesosoome, mis osalevad raku vaheseinte ehitamises, paljunemises ja on DNA kinnituskohaks. Bakterites toimub hingamine mesosoomides, sinivetikatel tsütoplasmaatilistes membraanides.
Paljudes bakterites ladestuvad raku sees varuained: polüsahhariidid, rasvad, polüfosfaadid. Varuained, mis sisalduvad ainevahetuses, võivad väliste energiaallikate puudumisel pikendada raku eluiga.
(1-raku sein, 2-välimine tsütoplasmaatiline membraan, 3-kromosoom (ringikujuline DNA molekul), 4-ribosoom, 5-mesosoom, 6-välise tsütoplasmaatilise membraani invaginatsioon, 7 vakuooli, 8-lippu, 9 virna membraanid, milles toimub fotosüntees)
Tavaliselt paljunevad bakterid kaheks jagunedes. Pärast raku pikenemist moodustub järk-järgult põiki vahesein, mis asetatakse väljastpoolt sissepoole, seejärel lahknevad tütarrakud või jäävad seotuks iseloomulike rühmadena - ketid, pakendid jne. Bakteri E. coli suurus kahekordistub iga 20 minuti järel.
Bakteritele on iseloomulik sporulatsioon. See algab tsütoplasma osa eraldumisega emarakust. Eraldunud osa sisaldab ühte genoomi ja on ümbritsetud tsütoplasmaatilise membraaniga. Seejärel kasvab eoste ümber rakusein, sageli mitmekihiline. Bakterites täheldatakse seksuaalset protsessi kahe raku vahelise geneetilise teabe vahetamise vormis. Seksuaalne protsess suurendab mikroorganismide pärilikku varieeruvust.
Enamik elusorganisme on ühendatud eukarüootide kuningriiki, sealhulgas taimede, seente ja loomade kuningriiki. eukarüootsed rakud on suuremad prokarüootsed rakud, koosnevad pinnaaparaadist, tuumast ja tsütoplasmast.
eukarüootne rakk
eukarüootne(eukarüootne)rakud sisaldavad tuuma raku elutegevuse koordineerimine, milles paikneb keha pärilik aparaat, ja arvukalt organellid erinevate funktsioonide täitmine. Enamik eukarüoote on aeroobid, see tähendab, et nad kasutavad energiavahetuseks õhuhapnikku.
Õppetund
raku organellid. Prokarüootsete ja eukarüootsete rakkude omadused "
(1. slaid)
Tunni eesmärk: rakkude püsikomponentide (organellide) struktuuride ja toimimise tunnuste tundmine; prokarüootsete ja eukarüootsete rakkude omaduste võrdlus
Varustus: multimeedia esitlused "Rakuorganoidid", "Prokarüootsed ja eukarüootsed rakud", bioloogia töövihik (11. klass), lk.61-64, jaotusmaterjal
Aja organiseerimine.
Tundide ajal:
Tunniplaan: (slaid 2 )
raku organellid
Mittemembraansed organellid
Membraani organellid
Prokarüootsed ja eukarüootsed rakud
Uue materjali õppimine:
raku organellid
Organellid (organellid) (slaid 3 ) on raku püsivad komponendid, mis täidavad selles spetsiifilisi funktsioone ning tagavad tema elutegevuse säilitamiseks vajalike protsesside ja omaduste elluviimise.
Organellidel võib olla nii membraani kui ka mittemembraanne struktuur.
Organellide klassifikatsioon (slaid 4) – Töö liigitusskeemi täitmisega: nad tuletavad meelde 9. klassis õpitud materjali (soovitav on märkmikusse salvestamine).
ÜLESANNE (väljatrükid igal laual): Täida tabel, kasutades õpetaja selgitusi ja õpiku materjale:
| Organoid | Struktuurne omadus | Nukleiinhapete olemasolu |
| Mittemembraansed organellid |
||
| Ribosoomid | ||
| Rakukeskus | ||
| mikrotuubulid | ||
| Mikrokiud | ||
| Kromosoomid | ||
| Ühemembraanilised organellid |
||
| Endoplasmaatiline retikulum | ||
| Golgi kompleks | ||
| Lüsosoomid | ||
| Topeltmembraanilised organellid |
||
| Mitokondrid | ||
| plastiidid | ||
Mittemembraansed organellid
RIBOSOOMID (slaid 5).
Ribosoom- elusraku kõige olulisem organell, sfääriline või kergelt ovaalne, 100-200 angströmi läbimõõduga, mis koosneb suurtest ja väikestest allüksustest (Slaid 6). Ribosoomide eesmärk on biosünteesida valke aminohapetest vastavalt antud mallile, mis põhineb messenger-RNA või mRNA edastatud geneetilisel teabel. Seda protsessi nimetatakse saade. Eukarüootsetes rakkudes paiknevad ribosoomid endoplasmaatilise retikulumi membraanidel, kuigi need võivad lokaliseerida ka lahtisel kujul tsütoplasmas. Tihti on ühe mRNA molekuliga seotud mitu ribosoomi, sellist struktuuri nimetatakse polüribosoom (Slaid 7 ) . Ribosoomide süntees eukarüootides toimub spetsiaalses tuumasiseses struktuuris - tuumas.
Eukarüootsed ribosoomid sisaldavad nelja rRNA molekuli
Esmakordselt kirjeldas ribosoome tihendatud osakeste või graanulitena Rumeenias sündinud rakubioloog George Palade 1950. aastate keskel. Termini "ribosoom" pakkus välja Richard Roberts 1958. aastal "mikrosomaalse fraktsiooni ribonukleoproteiini osakese" asemel.
RAKUKESKUS (TSENTROSOOM) (slaid 8).
Tsentrioolid on silindrilised valgustruktuurid, mis asuvad loomarakkude tuuma lähedal (taimedel tsentrioolid puuduvad). Tsentriool on silinder, mille külgpinna moodustavad üheksa mikrotuubulite komplekti. Mikrotuubulite arv komplektis võib erinevate organismide puhul varieeruda 1 kuni 3.
Tsentrioolide ümber on tsütoskeleti nn organiseerimiskeskus, piirkond, kuhu on rühmitatud raku mikrotuubulite miinusotsad.
Enne jagamist sisaldab rakk kahte tsentriooli, mis asuvad üksteise suhtes täisnurga all. Mitoosi ajal lahknevad nad raku erinevatesse otstesse, moodustades jagunemisspindli poolused. Pärast tsütokineesi saab iga tütarrakk ühe tsentriooli, mis kahekordistub järgmiseks jagunemiseks. Tsentrioolide kahekordistumine ei toimu mitte jagunemise, vaid uue struktuuri sünteesi teel, mis on risti olemasolevaga.
MIKROTUUBID (slaid 9)
Need on valkude intratsellulaarsed struktuurid, mis moodustavad tsütoskeleti.
Mikrotuubulid on 25 nm läbimõõduga silindrid, mille sees on õõnsus. Nende pikkus võib olla mõnest mikromeetrist kuni tõenäoliselt mõne millimeetrini närvirakkude aksonites. Mikrotuubulid on polaarsed, ühes otsas on isekoosnev ja teisest otsast lahtivõtmine. Rakkudes mängivad mikrotuubulid struktuurikomponentide rolli ja osalevad paljudes rakuprotsessides, sealhulgas mitoosis, tsütokineesis ja vesikulaarses transpordis.
Mikrotuubulite dünaamiline ebastabiilsus mängib olulist füsioloogilist rolli. Näiteks rakkude jagunemise ajal kasvavad mikrotuubulid väga kiiresti ja aitavad kaasa kromosoomide õigele orientatsioonile ja mitootilise spindli moodustumisele.
Rakus olevaid mikrotuubuleid kasutatakse osakeste transportimiseks "rööbastena". Membraani vesiikulid ja mitokondrid võivad liikuda mööda nende pinda. Transpordi läbi mikrotuubulite teostavad valgud, mida nimetatakse motoorseteks valkudeks.
MIKROFILAMENTID (Slaid 10 ).
Tsütoskeleti kontraktiilsed elemendid moodustuvad aktiini ja teiste kontraktiilsete valkude filamentidest. Osaleda raku tsütoskeleti moodustamises, amööbide liikumises jne. Nukleiinhapped puuduvad
KROMOSOOMID (slaid 11 ) - õpilased vastavad küsimusele, pidades meeles eelmise tunni materjali ja seejärel avaneb vastus slaidil.
Eukarüootide tuuma organellid, iga kromosoomi moodustavad üks DNA molekul ja valgumolekulid. Koosneb kahest niidist – kromatiididest, mis on ühendatud tsentromeeriga. Nad on geneetilise teabe kandjad.
Membraani organellid
Ühemembraanilised organellid
PLASMOLEMM (slaid 12 ) - õpilased vastavad küsimusele, meenutades eelmise tunni materjali ja seejärel avaneb vastus slaidil.
See on vedelik-mosaiikmudel, kus membraani lipiidikihid on valgumolekulidega läbi imbunud. See täidab rakuvälise keskkonna suhtes piiritlevat funktsiooni ja täidab transpordifunktsiooni. Nukleiinhappeid pole.
ENDOPLASMAATNE VÕRK (ER) (slaid 13)
Eukarüootses rakus on üksteisesse sisenevate membraanide (torude ja paakide) süsteem, mida nimetatakse endoplasmaatiliseks retikulumiks (või endoplasmaatiliseks retikulumiks, EPR või EPS). Seda ER osa, mille membraanide külge on kinnitatud ribosoomid, nimetatakse granuleeritud(või karm) (Klõpsake hiire nuppu) endoplasmaatiline retikulum, mille membraanidel toimub valgusüntees. Need sektsioonid, mille seintel ei ole ribosoome, klassifitseeritakse sile(või agranulaarne) EPR (Klõpsake hiirenuppu), osaleb lipiidide sünteesis. Sileda ja granuleeritud ER siseruumid ei ole isoleeritud, vaid lähevad üksteisesse ja suhtlevad tuumaümbrise valendikuga. Nukleiinhappeid pole.
GOLGI KOMPLEKS (PLATE COMPLEX)(Slaid 14 ) – vajutage hiirenuppu.
See on eukarüootse raku membraanstruktuur, mis on peamiselt ette nähtud endoplasmaatilises retikulumis sünteesitud ainete eritamiseks. (Slaid 15). Golgi kompleks sai nime Itaalia teadlase Camillo Golgi järgi, kes avastas selle esmakordselt 1898. slaid 16 ).
Golgi aparaadi mahutites küpsevad mõned granuleeritud ER membraanidel sünteesitud valgud, mis on mõeldud sekretsiooniks või lüsosoomide moodustamiseks. Golgi aparaat on asümmeetriline - paagid asuvad raku tuumale lähemal ( cis-Golgi) sisaldavad kõige vähem küpseid valke, nende paakide külge on pidevalt kinnitatud membraani vesiikulid - vesiikulid tärkab endoplasmaatilisest retikulumist. Ilmselt toimub samade vesiikulite abil küpsevate valkude edasine liikumine ühest paagist teise. Lõpuks organelli teisest otsast ( transs-Golgi) vesiikulid, mis sisaldavad täielikult küpseid valke, puhkevad.
LÜSOSOOM (Slaid 17 )
Need on membraanilised vesiikulid, mille suurus on kuni 2 mikronit. Lüsosoomid sisaldavad hüdrolüütilisi ensüüme, mis suudavad seedida valke, lipiide, süsivesikuid ja nukleiinhappeid. Lüsosoomid moodustuvad vesiikulitest, mis on eraldatud Golgi kompleksist, ja hüdrolüütilised ensüümid sünteesitakse eelnevalt jämedal en plasma retikulumiks.
Endotsüütiliste vesiikulitega ühinedes moodustuvad lüsosoomid seedetrakti vakuool (sekundaarne lüsosoom) , kus toimub orgaaniliste ainete lagunemine nende koostisesse kuuluvateks monomeerideks. Viimased sisenevad seedevakuooli membraani kaudu raku tsütoplasmasse. Täpselt nii juhtub, näiteks bakterite neutraliseerimine vererakkudes - neutrofiilid .
Sekundaarsed lüsosoomid, mille seedimisprotsess on lõppenud, praktiliselt ei sisalda ensüüme. Need sisaldavad ainult seedimata jääke.
Lüsosoomid osalevad ka rakumaterjalide, näiteks varutoitainete, aga ka funktsionaalse aktiivsuse kaotanud makromolekulide ja tervete organellide hävitamisel. (autofagia ). Raku patoloogiliste muutuste või selle vananemise korral võivad lüsosoomi membraanid hävida: ensüümid sisenevad tsütoplasmasse ja rakk ise seeditakse. - autolüüs . Mõnikord hävitatakse lüsosoomide abil terveid rakkude ja elundite komplekse. Näiteks kui kulles muutub konnaks, seedivad saba rakkudes olevad lüsosoomid seda: saba kaob ning selle protsessi käigus tekkinud ained imenduvad ja kasutavad ära teised keharakud.
VAKUULID
Need on suured membraansed vesiikulid või õõnsused tsütoplasmas, mis on täidetud rakumahlaga. Vakuoolid tekivad taimede ja seente rakkudes endoplasmaatilise retikulumi vesikulaarsetest jätketest või Golgi kompleksi vesiikulitest. Taimede meristemaatilistes rakkudes tekivad esmalt paljud väikesed vakuoolid. Kasvades nad ühinevad tsentraalne vakuool (Slaid 18) , mis hõivab kuni 70-90% raku mahust ja mida võivad läbida tsütoplasma ahelad.
Vakuoolide sisu rakumahl. See on erinevate anorgaaniliste ja orgaaniliste ainete vesilahus. Rakumahla keemiline koostis ja kontsentratsioon on väga varieeruvad ning sõltuvad taime tüübist, elundist, koest ja raku seisundist. Rakumahl sisaldab sooli, suhkruid (peamiselt sahharoosi, glükoosi, fruktoosi), orgaanilisi happeid (õun-, sidrun-, oksaal-, äädikhape jt), aminohappeid, valke. Need ained on ainevahetuse vaheproduktid, mis eemaldatakse ajutiselt raku ainevahetusest vakuooli. Nemad on tagavaraks raku ained.
Lisaks ainevahetuses taaskasutatavatele varuainetele sisaldab rakumahl fenoole, tanniine (tanniine), alkaloide, antotsüaniine, mis eemaldatakse ainevahetusest vakuooli ja isoleeritakse seega tsütoplasmast.
Tanniine leidub eriti sageli lehtede, koore, puidu, valmimata viljade ja seemnekestade rakkude mahlas (samuti tsütoplasmas ja membraanides). Alkaloide leidub näiteks kohviseemnetes (kofeiin), mooni viljades (morfiin) ja henbaanis (atropiin), lupiini vartes ja lehtedes (lupiin) jne. Arvatakse, et kokkutõmbava maitsega tanniinid, alkaloidid ja mürgised polüfenoolid toimivad. kaitsefunktsioon: nende mürgine (sageli mõru) maitse ja ebameeldiv lõhn tõrjuvad rohusööjaid, mis takistab nende söömist.
Sageli kogunevad vakuoolidesse ka rakkude elutegevuse lõpp-produktid. (jäätmed). Selline taimerakkude aine on kaltsiumoksalaat, mis ladestub vakuoolidesse erineva kujuga kristallidena.
Paljude taimede rakumahl sisaldab pigmente. , andes rakumahlale erinevaid värve. Pigmendid ja määrata lillede, viljade, pungade ja lehtede võrsete, samuti mõne taime (näiteks peedi) juurte värvus.
Mõnede taimede rakumahl sisaldab füsioloogiliselt aktiivseid aineid - fütohormoonid (kasvuregulaatorid), fütontsiidid, ensüümid . Viimasel juhul toimivad vakuoolid nagu lüsosoomid. Pärast rakusurma kaotab vakuooli membraan selektiivse läbilaskvuse ja sellest vabanevad ensüümid põhjustavad raku autolüüsi.
Tsentraalse vakuooli funktsioonid:
Toitainete, metaboliitide ja pigmentide kogunemine;
Ainevahetusproduktide eemaldamine tsütoplasmast;
Vee-soola ainevahetuse reguleerimine;
turgorurõhu säilitamine;
Osalemine makromolekulide ja rakustruktuuride hävitamises.
Seedetrakti vakuoolid (Slaid 19 ) loomarakud sisaldavad lüütilisi (lõhestavaid) ensüüme ja toiduosakesi. Siin toimub rakusisene seedimine.
algloomade eritavad vakuoolid sisaldavad vett ja selles lahustunud ainevahetusprodukte. Funktsioon - osmoregulatsioon, vedelate ainevahetusproduktide eemaldamine.
Topeltmembraanilised organellid
MITOKONDRIA (slaid 20)
Kahe membraaniga piklikud organellid. Need on rakkude energiajaamad. Mitokondrid on spetsiaalsed rakuorganellid, mille põhiülesanne on universaalse energiakandja ATP süntees. Hingamine (hapniku omastamine ja süsinikdioksiidi vabanemine) toimub ka mitokondrite ensümaatiliste süsteemide tõttu.
Mitokondritel on välimine membraan, mis koosneb kahest kihist, mida eraldab 60-80 angströmi pikkune ruum. Väljaulatuvad osad ulatuvad sisemisest kihist mitokondrite õõnsusse - cristae (vajutage hiirenuppu) . Cristae vaheline ruum on täidetud ainega, mida nimetatakse maatriks (vajutage hiirenuppu).
Maatriks sisaldab erinevaid ensüüme, mis osalevad hingamises ja ATP sünteesis. Sisemise mitokondriaalse membraani vesiniku potentsiaal on ATP sünteesi jaoks keskse tähtsusega. Sisaldab DNA-d ja RNA-d.
PLASTIIDID.
Plastiidid on eukarüootsete taimede ja mõnede fotosünteetiliste algloomade organellid. Kaetud topeltmembraaniga. Sisaldab DNA-d ja RNA-d. Moodustub plastiidide kogunemine rakus plastiid. Neid eristatakse värvi ja funktsiooni järgi kolm peamist plastiidide tüüpi(Slaid 21 ) :
Leukoplastid- plekimata plastiidid täidavad reeglina säilitusfunktsiooni. Kartulimugulate leukoplastides koguneb tärklis. Kõrgemate taimede leukoplastid võivad muutuda kloroplastideks või kromoplastideks.
Kromoplastid- kollaseks, punaseks või oranžiks värvitud plastiidid. Kromoplastide värvumine on seotud karotenoidide akumuleerumisega neis. Kromoplastid määravad sügislehtede, lillede kroonlehtede, juurviljade ja küpsete viljade värvi.
Kloroplastid- fotosünteetilisi pigmente kandvad plastiidid - klorofüllid. Rohelist värvi on neil kõrgematel taimedel, söel ja rohevetikatel. Fotosünteesis (ja vastavalt ka kloroplasti värvi määramisel) osalevate pigmentide kogum on erinevate taksonoomiliste jaotuste esindajatel erinev. Kloroplastidel on keeruline sisemine struktuur
Prokarüootsed ja eukarüootsed rakud
(kodutööna koos ülesande selgitusega tunnis)
Harjutus (slaid 22 ):
Vaatleme tabelit 2 lk 118
Täida töövihik lk 63-64
Täitke tabel, pannes märgid "+" ja "-"
| Rakkude struktuurid | prokarüootne rakk | eukarüootne rakk |
| raku sein | ||
| plasmalemma | ||
| Kromosoomid | ||
| Endoplasmaatiline retikulum | ||
| Golgi kompleks | ||
| Lüsosoomid | ||
| Mesosoom | ||
| Ribosoomid | ||
| Kaasamised |
Teabeallikad:
Gigani O.B. Üldbioloogia 9-11: Tabelid: skeemid / O. B. gigani. - M .: Humanitaarabi kirjastuskeskus VLADOS, 2007.
Kolman J., Rem K.-G. Visuaalne biokeemia: Per. temaga. - M.: Mir, 2000. http://yanko.lib.ru/books/biolog/nagl_biochem/04.htm
Vikipeedia – en.wikipedia.org
priroda.clow.ru/text/1190.htm - Entsüklopeedia "Taimed ja loomad"
biology.asvu.ru/page.php?id=17 –
www.college.ru/.../paragraph4/theory.html
shkola.lv/index.php?mode=lsntheme&themeid=104
Lisamaterjal õpetajale (Gigani O.B., 2007)
| Organoid | Struktuur | Funktsioonid | Nukleiinhapete olemasolu |
| Mittemembraansed organellid |
|||
| Ribosoomid | Moodustatud kahest allüksusest (suurest ja väikesest), mis on moodustatud rRNA ja valgu molekulidest | Valkude sünteesis osalemine | |
| Rakukeskus (tsentrosoom) | Koosneb kahest tsentrioolist, millest igaüks on õõnes silinder, mille moodustavad üheksa mikrotuubulite kolmikut. | Nad on osa raku mitootilisest aparaadist, osalevad raku jagunemises | |
| mikrotuubulid | Õõnes silindrilised struktuurid | Nad moodustavad raku tsütoskeleti, spindli, tsentrioolid, lipud ja ripsmed | |
| Mikrokiud | Tsütoskeleti kontraktiilsed elemendid, mis on moodustatud aktiini ja teiste kontraktiilsete valkude filamentidest | Osalemine raku tsütoskeleti moodustamises, amööboidne liikumine, endotsütoos, tsüklos | |
| Kromosoomid | Eukarüootsete rakkude tuuma organellid, iga kromosoomi moodustavad üks DNA molekul ja valgumolekulid | Geneetilise informatsiooni kandjad | |
| Ühemembraanilised organellid |
|||
| Plasmolemma (tsütolemma) Endoplasmaatiline retikulum Sile (agranulaarne) EPS Kare (granuleeritud) EPS | Elementaarne membraan, mis katab raku välispinna Membraanide süsteem, mis moodustab tuubulid, vesiikulid, tsisternid, tuubulid. Ühendatud plasmamembraani ja tuumamembraaniga. Membraanide pinnal on ensüümid, mis katalüüsivad lipiidide ja süsivesikute sünteesi. Ribosoomid asuvad membraanide pinnal. | Raku kuju säilitamine, kaitse ebasoodsate välismõjude eest, ainete transport rakku ja sealt välja, retseptor (tajub membraani sisse ehitatud erinevatele molekulidele, tajub keskkonnasignaale) Ainete transport rakus, raku jagunemine sektsioonideks, valkude translatsioonijärgne modifitseerimine. Lipiidide ja süsivesikute süntees, toksiliste ainete kogunemine ja eemaldamine Valkude süntees membraaniga seotud ribosoomidel, mis on ühendatud kompleksideks - polüsoomideks | |
| Golgi kompleks (lamellkompleks) | Erinevate organismide rakkude struktuur on väga erinev. Golgi kompleksi struktuurne ja funktsionaalne üksus - diktüosoom - 5-20 lamedat tsisterni virn, mis lõpevad torukeste ja vesiikulite võrgustikuga | Ainete muutmine; nende pakkimine membraani vesiikulitesse, mida seejärel rakk kasutab või sealt eemaldatakse; teatud ainete süntees; rakumembraanide moodustumine; lüsosoomide moodustumine | |
| Lüsosoomid | Membraani vesiikulid on ümarad ja sisaldavad lüütiline (jagamine) ensüümid | Osalemine seedevakuoolide moodustamises (rakusisene seedimine); suurte rakumolekulide hävitamine; üksikute rakustruktuuride (autolüüs) ja kogu raku lüüs (hävitamine); ajutiste volituste kaotamine | |
| Taimeraku keskvakuool Loomarakkude seedevakuoolid algloomade eritavad vakuoolid | Membraaniga ümbritsetud õõnsused, mis sisaldavad erinevate lahustunud ainetega vesilahust. Seda piirab tonoplast - membraan. Täidetud rakumahlaga (lahustunud orgaanilised ja anorgaanilised ained, pigmendid, metaboliidid). Moodustati EPS-i osalusel. | Toitainete, metaboliitide ja pigmentide kogunemine; ainevahetusproduktide eemaldamine tsütoplasmast; vee-soola ainevahetuse reguleerimine; turgorirõhu säilitamine; osalemine makromolekulide ja rakustruktuuride hävitamises. rakusisene seedimine Osmoregulatsioon, vedelate ainevahetusproduktide eemaldamine | |
| Topeltmembraanilised organellid |
|||
| Mitokondrid | Välimine membraan on sile, sisemine moodustab väljakasvu - cristae. Sees on maatriks - poolvedel aine, mis sisaldab ensüüme, ringikujulisi DNA molekule, RNA molekule, ribosoome | ATP süntees | |
| plastiidid Protoplastiidid Kloroplastid Kromoplastid Leukoplastid | Välismembraan on sile, sisemine membraan on sukeldatud stroomas - poolvedelas aines. Sisaldab ringikujulisi DNA molekule, RNA molekule ja ribosoome Ei oma värvust Sisemembraan moodustab lamestatud kotikesed - tülakoidid, milles paiknevad pigmendimolekulid (klorofüll, karotenoidid), tülakoidide rühm moodustab grana Sisemembraan moodustab mõned tülakoidid | Plastiidid, millest moodustuvad igat tüüpi plastiidid (kloroplastid, leukoplastid, kromoplastid) Fotosüntees, võib muutuda kromoplastideks Lille kroonlehtede, viljade, lehtede, vahel ka juurte värvimine Tärklise, õli, valkude süntees ja akumuleerumine võib muutuda kloroplastideks ja kromoplastideks | |
Prokarüootsete ja eukarüootsete rakkude võrdlevad omadused
| Rakkude struktuurid | prokarüootne rakk | eukarüootne rakk |
| raku sein | Taime- ja seenerakkudes |
|
| plasmalemma | ||
| Kromosoomid | - (seal on nukleotiid - 1 ringikujuline DNA molekul) | |
| Endoplasmaatiline retikulum | ||
| Golgi kompleks | ||
| Lüsosoomid | ||
| Kahe membraaniga organellid (plastiidid, mitokondrid) | ||
| Mesosoom | ||
| Ribosoomid | ||
| Kaasamised |
Raku ehituse järgi jagunevad elusorganismid prokarüootid ja eukarüoot. Mõlema rakud on ümbritsetud plasmamembraan, millest väljaspool paljudel juhtudel on raku sein. Raku sees on poolvedel tsütoplasma. Prokarüootsed rakud on aga palju lihtsamad kui eukarüootsed rakud.
Põhiline geneetiline materjal prokarüootid (kreeka keelest. umbes- enne ja karyon- tuum) asub tsütoplasmas ringikujulise DNA molekulina. See molekul ( nukleoid) ei ole ümbritsetud eukarüootidele iseloomuliku tuumamembraaniga ja on kinnitunud plasmamembraanile (joonis 1). Seega ei ole prokarüootidel hästi moodustunud tuum. Lisaks nukleoidile leidub prokarüootses rakus sageli ka väike ringikujuline DNA molekul, nn. plasmiid. Plasmiidid võivad liikuda ühest rakust teise ja integreeruda DNA põhimolekuli.
Mõnedel prokarüootidel on plasmamembraani väljakasvud: mesosoomid, lamellaarsed tülakoidid, kromatofoorid. Need sisaldavad ensüüme, mis osalevad fotosünteesis ja hingamises. Lisaks on mesosoomid seotud DNA sünteesi ja valkude sekretsiooniga.
Prokarüootsed rakud on väikesed, nende läbimõõt on 0,3-5 mikronit. Kõigi prokarüootide (välja arvatud mükoplasmad) plasmamembraani välisküljel on raku sein. See koosneb kihtidena laotud valkude ja oligosahhariidide kompleksidest, kaitseb rakku ja säilitab selle kuju. See on plasmamembraanist eraldatud väikese membraanidevahelise ruumiga.
Prokarüootide tsütoplasmas leidub ainult mittemembraanseid organelle. ribosoomid. Prokarüootide ja eukarüootide ribosoomide struktuur on sarnane, kuid prokarüootide ribosoomid on väiksemad ja ei ole membraani külge kinnitatud, vaid paiknevad otse tsütoplasmas.

Paljud prokarüootid on liikuvad ja võivad lipukeste abil ujuda või libiseda.
Prokarüootid paljunevad tavaliselt lõhustumise teel kaheks ( binaarne). Jagunemisele eelneb väga lühike kromosoomide kahekordistumise ehk replikatsiooni etapp. Seega on prokarüootid haploidsed organismid.
Prokarüootide hulka kuuluvad bakterid ja sinivetikad ehk tsüanobakterid. Prokarüootid ilmusid Maale umbes 3,5 miljardit aastat tagasi ja olid tõenäoliselt esimene rakuline eluvorm, millest said alguse tänapäevased prokarüootid ja eukarüootid.
eukarüootid (kreeka keelest. eu- tõsi, karyon- tuum), erinevalt prokarüootidest on moodustunud tuum, mida ümbritseb tuumaümbris- kahekihiline membraan. Tuumas leiduvad DNA molekulid ei ole suletud (lineaarsed molekulid). Lisaks tuumale sisaldub osa geneetilisest informatsioonist mitokondrite ja kloroplastide DNA-s. Eukarüootid ilmusid Maale umbes 1,5 miljardit aastat tagasi.
Erinevalt prokarüootidest, mida esindavad üksikud organismid ja koloniaalvormid, võivad eukarüootid olla üherakulised (näiteks amööb), koloniaalsed (volvox) ja mitmerakulised. Need on jagatud kolmeks suureks kuningriigiks: loomad, taimed ja seened.
Eukarüootsete rakkude läbimõõt on 5–80 µm. Nagu prokarüootsed rakud, on eukarüootsed rakud ümbritsetud plasmamembraan koosneb valkudest ja lipiididest. See membraan toimib selektiivse barjäärina, mis on mõnele ühendile läbilaskev ja teistele mitteläbilaskev. Väljaspool plasmamembraani on tugev raku sein, mis taimedes koosneb peamiselt tselluloosikiududest ja seentes - kitiinist. Rakuseina põhiülesanne on tagada rakkude püsiv kuju. Kuna plasmamembraan on vett läbilaskev ning taime- ja seenerakud puutuvad tavaliselt kokku lahustega, mille ioontugevus on väiksem kui rakus oleva lahuse ioontugevus, siis satub vesi rakkudesse. Tänu sellele suureneb rakkude maht, plasmamembraan hakkab venima ja võib puruneda. Rakusein takistab rakkude laienemist ja hävimist.
Loomadel rakusein puudub, kuid plasmamembraani välimine kiht on rikastatud süsivesikute komponentidega. Seda loomarakkude plasmamembraani välimist kihti nimetatakse glükokalüks. Mitmerakuliste loomade rakud ei vaja tugevat rakuseina, kuna on ka teisi mehhanisme, mis tagavad rakumahu reguleerimise. Kuna meres elavate paljurakuliste loomade ja ainuraksete organismide rakud on keskkonnas, kus ioonide summaarne kontsentratsioon on lähedane ioonide rakusisesele kontsentratsioonile, siis rakud ei paisu ega lõhke. Magedas vees elavatel üherakulistel loomadel (amööb, ripslik king) on kokkutõmbuvad vakuoolid, mis toovad rakku siseneva vee pidevalt välja.
Eukarüootse raku struktuurikomponendid
Plasmamembraani all oleva raku sees on tsütoplasma. Tsütoplasma (hüaloplasma) põhiaine on anorgaaniliste ja orgaaniliste ühendite kontsentreeritud lahus, mille põhikomponendid on valgud. See on kolloidne süsteem, mis võib muutuda vedelast olekust geeliks ja vastupidi. Märkimisväärne osa tsütoplasma valkudest on ensüümid, mis viivad läbi erinevaid keemilisi reaktsioone. asub hüaloplasmas organellid, täidab rakus erinevaid funktsioone. Organellid võivad olla membraanilised (tuum, Golgi aparaat, endoplasmaatiline retikulum, lüsosoomid, mitokondrid, kloroplastid) ja mittemembraansed (rakukeskus, ribosoomid, tsütoskelett).
Membraani organellidmembraani organellide põhikomponent on membraan. Bioloogilised membraanid on ehitatud üldpõhimõtte järgi, kuid erinevate organellide membraanide keemiline koostis on erinev. Kõik rakumembraanid on õhukesed kiled (paksus 7–10 nm), mis põhinevad lipiidide topeltkihil (kakskihil), mis on paigutatud nii, et molekulide laetud hüdrofiilsed osad puutuvad kokku keskkonnaga ja hüdrofoobsed rasvhappejäägid. iga monokiht on suunatud membraani sisse ja puudutavad üksteist.sõbraga (joonis 3). Valgumolekulid (integraalsed membraanivalgud) on ehitatud lipiidide kaksikkihti selliselt, et valgumolekuli hüdrofoobsed osad puutuvad kokku lipiidimolekulide rasvhappejääkidega ning hüdrofiilsed osad puutuvad kokku keskkonnaga. Lisaks on osa lahustuvatest (mittemembraansed valgud) membraaniga seotud peamiselt ioonsete interaktsioonide tõttu (perifeerse membraani valgud). Membraanide koostises on paljude valkude ja lipiidide külge kinnitatud ka süsivesikute fragmendid. Seega on bioloogilised membraanid lipiidkiled, millesse on integreeritud valgud.
Membraanide üks põhifunktsioone on piiri loomine raku ja keskkonna ning raku erinevate sektsioonide vahel. Lipiidide kaksikkiht on läbilaskev peamiselt rasvlahustuvate ühendite ja gaaside jaoks, hüdrofiilsed ained transporditakse läbi membraanide spetsiaalsete mehhanismide abil: madala molekulmassiga - kasutades erinevaid kandjaid (kanalid, pumbad jne) ja suure molekulmassiga - kasutades protsesse. ekso- ja endotsütoos(joonis 4).

Riis. 4. Ainete membraani läbimise skeem
Kell endotsütoos teatud ained sorbeeritakse membraani pinnal (koostoime tõttu membraanivalkudega). Selles kohas moodustub membraani invaginatsioon tsütoplasmasse. Seejärel eraldatakse membraanist vesiikul, mille sees on ülekantud ühend. Sellel viisil, endotsütoos- see on väliskeskkonna kõrgmolekulaarsete ühendite ülekandmine rakku, mida ümbritseb membraani osa. Vastupidine protsess on eksotsütoos on ainete transport rakust väljapoole. See tekib transporditavate kõrgmolekulaarsete ühenditega täidetud mulli plasmamembraaniga sulandumisel. Vesiikulite membraan sulandub plasmamembraaniga ja selle sisu valatakse välja.
Kanalid, pumbad ja muud transporterid on integreeritud membraanivalgu molekulid, mis tavaliselt moodustavad membraanis poori.
Lisaks ruumi jagamise ja selektiivse läbilaskvuse tagamise funktsioonidele on membraanid võimelised signaale tajuma. Seda funktsiooni täidavad retseptorvalgud, mis seovad signaalmolekule. Üksikud membraanivalgud on ensüümid, mis viivad läbi teatud keemilisi reaktsioone.
Tuum - suur raku organell, mis on ümbritsetud tuumamembraaniga ja millel on tavaliselt sfääriline kuju. Rakus on ainult üks tuum ja kuigi on olemas mitmetuumalisi rakke (skeletilihasrakud, mõned seened) või ilma tuumata (imetajate erütrotsüüdid ja vereliistakud), tekivad need rakud mononukleaarsetest eellasrakkudest.
Kerneli põhifunktsioon on geneetilise teabe säilitamine, ülekandmine ja müük. Siin dubleeritakse DNA molekulid, mille tulemusena saavad tütarrakud jagunemisel sama geneetilise materjali. Tuumas, kasutades maatriksina DNA molekulide (geenide) üksikuid sektsioone, sünteesitakse RNA molekulid: valgusünteesiks vajalikud informatsioonilised (mRNA), transpordi (tRNA) ja ribosomaalsed (rRNA) molekulid. Tuumas koostatakse rRNA molekulidest ja valkudest ribosoomi subühikud, mis sünteesitakse tsütoplasmas ja kanduvad tuuma.
Tuum koosneb tuumamembraanist, kromatiinist (kromosoomid), tuumast ja nukleoplasmast (karüoplasmast).

Riis. 5. Kromatiini struktuur: 1 - nukleosoom, 2 - DNA
Mikroskoobi all on tuuma sees nähtavad tiheda aine tsoonid - kromatiin. Mittejagunevates rakkudes täidab see ühtlaselt tuuma ruumala või kondenseerub eraldi kohtadesse tihedamate piirkondadena ja määrib hästi põhivärvidega. Kromatiin on DNA ja valkude kompleks (joonis 5), enamasti positiivselt laetud histoonid.
DNA molekulide arv tuumas on võrdne kromosoomide arvuga. Kromosoomide arv ja kuju on liigi ainulaadne omadus. Iga kromosoom sisaldab ühte DNA molekuli, mis koosneb kahest omavahel ühendatud ahelast ja on 2 nm paksuse kaksikheeliksi kujuga. Selle pikkus ületab oluliselt raku läbimõõtu: see võib ulatuda mitme sentimeetrini. DNA molekul on negatiivselt laetud, seega saab see voltida (kondenseeruda) alles pärast seondumist positiivselt laetud histooni valkudega (joonis 6).
Esiteks keerdub DNA topeltahel ümber üksikute histoonide plokkide, millest igaüks sisaldab 8 valgumolekuli, moodustades umbes 10 nm paksuste "helmeste nööril" kujul struktuuri. Helmeid nimetatakse nukleosoomideks. Nukleosoomide moodustumise tulemusena väheneb DNA molekuli pikkus umbes 7 korda. Järgmisena volditakse nukleosoomidega niit, moodustades umbes 30 nm paksuse trossi kujul oleva struktuuri. Seejärel kinnitatakse selline silmuste kujul painutatud köis valkude külge, mis moodustavad kromosoomi aluse. Selle tulemusena moodustub struktuur paksusega umbes 300 nm. Selle struktuuri edasine kondenseerumine viib kromosoomi moodustumiseni.
Jagunemise vahel rullub kromosoom osaliselt lahti. Selle tulemusena vabanevad DNA molekuli üksikud lõigud, mis peaksid antud rakus ekspresseeruma, valkudest ja venitada, mis võimaldab RNA molekule sünteesides neist infot lugeda.
Tuum on teatud tüüpi matriitsi DNA, mis vastutab rRNA sünteesi eest ja on kokku pandud tuuma erinevatesse piirkondadesse. Tuum on tuuma kõige tihedam struktuur, see ei ole eraldi organell, vaid on üks kromosoomi lookusi. See toodab rRNA-d, mis seejärel moodustab valkudega kompleksi, moodustades ribosomaalsed subühikud, mis lähevad tsütoplasmasse.
Tuuma mittehistoonvalgud moodustavad tuuma sees struktuurse võrgu. Seda esindab tuumaümbrise all olev fibrillide kiht. Selle külge on kinnitatud fibrillide tuumasisene võrgustik, mille külge on kinnitatud kromatiini fibrillid.
Tuumaümbris koosneb kahest membraanist: välimisest ja sisemisest, mis on eraldatud membraanidevahelise ruumiga. Välismembraan on kontaktis tsütoplasmaga, see võib sisaldada polüribosoome ja see võib ise siseneda endoplasmaatilise retikulumi membraanidesse. Sisemembraan on seotud kromatiiniga. Seega tagab tuumaümbris kromosomaalse materjali fikseerimise tuuma kolmemõõtmelises ruumis.
Tuuma kestal on ümmargused augud - tuumapoorid(joonis 7). Pooride piirkonnas sulguvad välimine ja sisemine membraan ning moodustuvad fibrillide ja graanulitega täidetud augud. Poori sees on keeruline valkude süsteem, mis tagab makromolekulide selektiivse sidumise ja ülekande. Tuumapooride arv sõltub raku ainevahetuse intensiivsusest.
Endoplasmaatiline retikulum, või endoplasmaatiline retikulum (EPR) on veider kanalite, vakuoolide, lamestatud kotikeste võrgustik, mis on omavahel ühendatud ja eraldatud hüaloplasmast membraaniga (joonis 8).
Eristama karm ja sile EPR . Karedate ER membraanidel on ribosoomid(joonis 9), mis sünteesivad rakust eritunud või plasmamembraani inkorporeeritud valke. Äsja sünteesitud valk lahkub ribosoomist ja liigub spetsiaalse kanali kaudu endoplasmaatilise retikulumi õõnsusse, kus toimub translatsioonijärgne modifikatsioon, näiteks seondumine süsivesikutega, polüpeptiidahela osa proteolüütiline lõhustamine ja S moodustumine. – S-sidemed ahela tsüsteiinijääkide vahel. Lisaks transporditakse need valgud Golgi kompleksi, kus need on kas osa lüsosoomidest või sekretoorsetest graanulitest. Mõlemal juhul on need valgud membraani vesiikuli (vesiikuli) sees.

Riis. Joonis 9. Valgusünteesi skeem töötlemata ER-s: 1 – väike ja
2 - ribosoomi suur subühik; 3 – rRNA molekul;
4 - töötlemata EPR; 5 - äsja sünteesitud valk
Siledal ER-l puuduvad ribosoomid. Selle põhiülesanne on lipiidide süntees ja süsivesikute ainevahetus. See on hästi arenenud näiteks neerupealiste koore rakkudes, mis sisaldavad steroidhormoonide sünteesi tagavaid ensüüme. Maksarakkude sile ER sisaldab ensüüme, mis oksüdeerivad (detoksifitseerivad) organismile võõraid hüdrofoobseid ühendeid, näiteks ravimeid.

Riis. 10. Golgi aparaat: 1 - vesiikulid; 2 - tankid
Golgi kompleks (joon. 10) koosneb 5–10 paralleelselt asetsevast lamedast membraaniga seotud õõnsusest. Nende kettakujuliste konstruktsioonide otsaosad on pikendustega. Selliseid moodustisi võib rakus olla mitu. Golgi kompleksi tsoonis on suur hulk membraani vesiikuleid. Mõned neist on põhistruktuuri otsaosadest kinni tõmmatud sekretoorsete graanulite ja lüsosoomide kujul. Mõned väikesed vesiikulid (vesiikulid), mis kannavad karedas ER-s sünteesitud valke, liiguvad Golgi kompleksi ja ühinevad sellega. Seega on Golgi kompleks seotud töötlemata EPR-is sünteesitud toodete kogumise ja edasise muutmisega ning nende sorteerimisega.
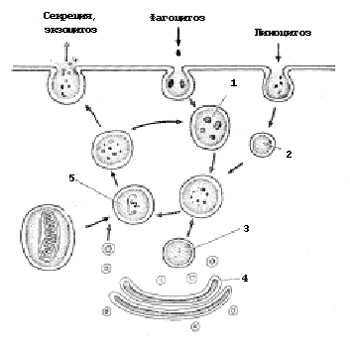
Riis. 11. Lüsosoomide teke ja funktsioonid: 1 - fagosoom; 2 - pinotsüütiline vesiikul; 3 - primaarne lüsosoom; 4 - Golgi aparaat; 5 - sekundaarne lüsosoom
Lüsosoomid - need on vakuoolid (joonis 11), mis on piiratud ühe membraaniga ja mis tärkavad Golgi kompleksist. Lüsosoomide sees on üsna happeline keskkond (pH 4,9–5,2). On hüdrolüütilisi ensüüme, mis lõhustavad happelise pH juures erinevaid polümeere (proteaasid, nukleaasid, glükosidaasid, fosfataasid, lipaasid). Need primaarsed lüsosoomid sulanduvad endotsüütiliste vakuoolidega, mis sisaldavad lõhustatavaid komponente. Ained, mis sisenevad sekundaarsesse lüsosoomi, lagunevad monomeerideks ja kantakse läbi lüsosoomi membraani hüaloplasmasse. Seega osalevad lüsosoomid rakusisese seedimise protsessides.
Mitokondrid ümbritsetud kahe membraaniga: välimine, mis eraldab mitokondrid hüaloplasmast, ja sisemine, mis piiritleb selle sisemise sisu. Nende vahele jääb 10–20 nm laiune membraanidevaheline ruum. Sisemembraan moodustab arvukalt väljakasvu ( cristae). See membraan sisaldab ensüüme, mis tagavad väljaspool mitokondreid moodustunud aminohapete, suhkrute, glütserooli ja rasvhapete oksüdatsiooni (Krebsi tsükkel) ning teostavad elektronide ülekande hingamisahelas (skeem). Tänu elektronide ülekandele mööda hingamisahelat kõrgelt energiatasemelt madalamale salvestatakse osa vabanenud vabast energiast ATP-na – raku universaalse energiavaluutana. Seega on mitokondrite põhiülesanne erinevate substraatide oksüdeerimine ja ATP molekulide süntees.
Kahe elektroni ülekande skeem mööda hingamisahelat
Mitokondrite sees on ümmargune DNA molekul, mis kodeerib mõningaid mitokondriaalseid valke. Mitokondrite siseruumis (maatriksis) on prokarüootidega sarnased ribosoomid, mis tagavad nende valkude sünteesi.
Asjaolu, et mitokondritel on oma ümmargune DNA ja prokarüootsed ribosoomid, on viinud hüpoteesini, et mitokondrid on iidse prokarüootse raku järeltulija, mis kunagi sattus eukarüootsesse rakku ja võtsid evolutsiooni käigus oma funktsioonid.
Riis. 12. Kloroplastid (A) ja tülakoidmembraanid (B)
plastiidid - taimeraku organellid, mis sisaldavad pigmente. AT kloroplastid Sisaldab klorofülli ja karotenoide kromoplastid- karotenoidid, leukoplastid pigmente pole. Plastiidid on ümbritsetud topeltmembraaniga. Nende sees on lamedate vesiikulite kujul membraanide süsteem, mida nimetatakse tülakoidid(joonis 12). Tülakoidid on laotud virnadesse, mis meenutavad plaadivirnasid. Pigmendid on põimitud tülakoidmembraanidesse. Nende põhiülesanne on valguse neeldumine, mille energia tülakoidmembraani sisse ehitatud ensüümide abil muudetakse tülakoidmembraanil H + ioonide gradiendiks. Nagu mitokondritel, on ka plastiididel oma ringikujuline DNA ja prokarüootset tüüpi ribosoomid. Ilmselt on plastiidid ka eukarüootsete rakkudega sümbioosis elav prokarüootne organism.
Ribosoomid on mittemembraansed rakulised organellid, mida leidub nii pro- kui ka eukarüootsetes rakkudes. Eukarüootsed ribosoomid on suuremad kui prokarüootsed, nende suurus on 25x20x20 nm. Ribosoom koosneb üksteisega külgnevatest suurtest ja väikestest subühikutest. MRNA ahel paikneb toimivas ribosoomi allüksuste vahel.
Iga ribosoomi subühik on ehitatud rRNA-st, tihedalt pakitud ja seotud valkudega. Ribosoomid võivad tsütoplasmas vabalt paikneda või olla seotud ER-membraanidega. Vabad ribosoomid võivad olla üksikud, kuid võivad moodustada polüsoome, kui mitu ribosoomi paikneb järjestikku ühel mRNA ahelal. Ribosoomide põhiülesanne on valkude süntees.
tsütoskelett - see on raku luu-lihassüsteem, sealhulgas valkfilamentsed (fibrillaarsed) moodustised, mis on raku skelett ja täidavad motoorset funktsiooni. Tsütoskeleti struktuurid on dünaamilised, tekivad ja lagunevad. Tsütoskeleti esindavad kolme tüüpi moodustised: vahepealsed filamendid(kiud läbimõõduga 10 nm), mikrofilament s (kiud läbimõõduga 5–7 nm) ja mikrotuubulid. Vahefilamendid on hargnemata valgustruktuurid filamentide kujul, mis on sageli paigutatud kimpudesse. Nende valgu koostis on erinevates kudedes erinev: epiteelis koosnevad nad keratiinist, fibroblastides - vimentiinist, lihasrakkudes - desmiinist. Vahefilamendid täidavad tugiraami funktsiooni.
Mikrokiud - need on fibrillaarsed struktuurid, mis paiknevad kimpude või kihtidena otse plasmamembraani all. Need on selgelt nähtavad amööbi prolegides, fibroblastide liikuvates protsessides ja sooleepiteeli mikrovillides (joonis 13). Mikrofilamendid koosnevad kontraktiilsetest valkudest aktiinist ja müosiinist ning on intratsellulaarne kontraktiilne aparaat.
mikrotuubulid on osa raku ajutistest ja püsivatest struktuuridest. Ajaliste hulka kuuluvad jagunemisspindli, jagunemistevahelise rakkude tsütoskeleti elemendid ning püsivate hulka kuuluvad rakukeskuse ripsmed, lipud ja tsentrioolid. Mikrotuubulid on sirged õõnsad silindrid läbimõõduga umbes 24 nm, nende seinad on moodustatud ümarate tubuliini valgu molekulidest. Elektronmikroskoobi all on näha, et mikrotuubuli ristlõike moodustavad 13 rõngasse ühendatud alaühikut. Mikrotuubulid on kõigi eukarüootsete rakkude hüaloplasmas. Mikrotuubulite üks ülesandeid on luua rakkude sees karkass. Lisaks liiguvad väikesed vesiikulid mööda mikrotuubuleid, justkui rööbastel.
Rakukeskus koosneb kahest teineteise suhtes täisnurga all paiknevast tsentrioolist ja nendega seotud mikrotuubulitest. Need jagunevate rakkude organellid osalevad jagunemisspindli moodustamises. Tsentriooli aluseks on 9 ümbermõõtu paiknevat mikrotuubulite kolmikut, mis moodustavad õõnsa silindri, mille laius on 0,2 µm ja pikkus 0,3–0,5 µm. Rakkude jagunemiseks valmistudes tsentrioolid eralduvad ja kahekordistuvad. Enne mitoosi osalevad tsentrioolid spindli mikrotuubulite moodustumisel. Kõrgematel taimerakkudel ei ole tsentrioole, kuid neil on sarnane mikrotuubulite organiseerimise keskus.