אחד התהליכים החשובים ביותר בהתפתחות האישית של אורגניזם חי הוא מיטוזה. במאמר זה ננסה להסביר בקצרה ובבהירות אילו תהליכים מתרחשים במהלך חלוקת התא, ונדבר על המשמעות הביולוגית של מיטוזה.
הגדרת מושג
מספרי הלימוד בביולוגיה לכיתה י', אנו יודעים שמיטוזה היא חלוקת תאים, וכתוצאה מכך נוצרים מתא אם אחד שני תאי בת עם אותה מערכת כרומוזומים.
בתרגום מהשפה היוונית העתיקה, המונח "מיטוזה" פירושו "חוט". זה כמו קישור בין תאים ישנים לחדשים, שבו מאוחסן הקוד הגנטי.
תהליך החלוקה בכללותו מתחיל מהגרעין ומסתיים בציטופלזמה. זה מכונה מחזור מיטוטי, המורכב מהשלב של מיטוזה ואינטרפאזה. כתוצאה מחלוקה של תא סומטי דיפלואידי, נוצרים שני תאי בת. עקב תהליך זה, מתרחשת עלייה במספר תאי הרקמה.
שלבים של מיטוזה
בהתבסס על מאפיינים מורפולוגיים, תהליך החלוקה מחולק לשלבים הבאים:
- פרופאזה ;
בשלב זה, הגרעין מתעבה, הכרומטין מתעבה בתוכו, שמתפתל לספירלה, צופים בכרומוזומים במיקרוסקופ.
4 המאמרים המוביליםשקרא יחד עם זה
בהשפעת אנזימים, הגרעינים והממברנות שלהם מתמוססים, הכרומוזומים בתקופה זו מסודרים באופן אקראי בציטופלזמה. מאוחר יותר, מתרחשת הפרדה של centrioles לקטבים, נוצר ציר של חלוקת תאים, החוטים שלו מחוברים לקטבים ולכרומוזומים.
שלב זה מאופיין בהכפלת DNA, אך זוגות כרומוזומים עדיין מחזיקים אחד בשני.
לפני שלב הפרופזה יש לתא הצמח שלב הכנה - פרפרופאז. מהי ההכנה של התא למיטוזה ניתן להבין בשלב זה. הוא מאופיין ביצירת טבעת קדם-פרופאזה, פרגמוזומים ויצירת גרעין של מיקרוטובולים סביב הגרעין.
- פרומטפאזה ;
בשלב זה, הכרומוזומים מתחילים לנוע ופונים לכיוון הקוטב הקרוב.
בספרי לימוד רבים, קדם-פרופאזה ופרומטופאז מכונים שלב הפרופזה.
- מטאפאזה ;
על שלב ראשוניהכרומוזומים ממוקמים בחלק המשווני של הציר, כך שלחץ הקטבים פועל עליהם באופן שווה. במהלך שלב זה, מספר המיקרוטובולים בציר גדל ומתחדש כל הזמן.
הכרומוזומים מסתדרים בזוגות בספירלה לאורך קו המשווה של הציר בסדר קפדני. הכרומטידות מתנתקות בהדרגה, אך עדיין נאחזות בחוטי הציר.
- אנפאזה ;
בשלב זה מתרחשת התארכות של הכרומטידות, שמתפצלות בהדרגה לכיוון הקטבים, כאשר חוטי הציר מתכווצים. נוצרים כרומוזומי בת.
מבחינת זמן, זה השלב הקצר ביותר. כרומטידות אחיות נפרדות לפתע ועוברות לקטבים שונים.
- טלופאז ;
זהו השלב האחרון של החלוקה כאשר הכרומוזומים מתארכים ונוצרת מעטפת גרעינית חדשה ליד כל קוטב. החוטים שהרכיבו את הציר נהרסים לחלוטין. בשלב זה, הציטופלזמה מתחלקת.
השלמת השלב האחרון עולה בקנה אחד עם חלוקת תא האם, הנקראת ציטוקינזיס. זה תלוי במעבר של תהליך זה כמה תאים נוצרים במהלך החלוקה, יכולים להיות שניים או יותר.
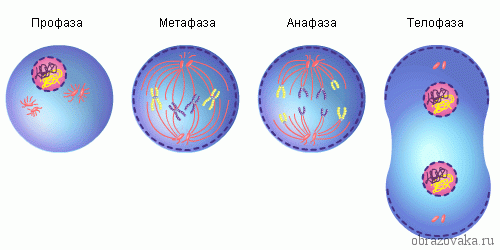
אורז. 1. שלבי מיטוזה
המשמעות של מיטוזיס
אין להכחיש את המשמעות הביולוגית של תהליך חלוקת התא.
- בזכותו אפשר לשמור על מערך קבוע של כרומוזומים.
- רבייה של תא זהה אפשרית רק על ידי מיטוזה. בדרך זו מחליפים תאי עור, אפיתל מעי, תאי דם אדומים שמחזור החיים שלהם הוא 4 חודשים בלבד.
- העתקה, ומכאן שימור המידע הגנטי.
- הבטחת התפתחות וצמיחה של תאים, שבגללם נוצר אורגניזם רב תאי מזיגוטה חד-תאית.
- בעזרת חלוקה כזו מתאפשרת התחדשות של חלקי גוף בכמה אורגניזמים חיים. לדוגמה, קרני כוכב הים משוחזרות.

אורז. 2. התחדשות כוכבי הים
- הבטחת רבייה א-מינית. לדוגמה, ניצני הידרה, כמו גם ריבוי וגטטיבי של צמחים.

אורז. 3. הידרה באדינג
מה למדנו?
חלוקת תאים נקראת מיטוזה. הודות לו, המידע הגנטי של התא מועתק ונשמר. התהליך מתרחש במספר שלבים: שלב הכנה, פרופאזה, מטאפאזה, אנפאזה, טלופזה. כתוצאה מכך נוצרים שני תאי בת, הדומים לחלוטין לתא האם המקורי. בטבע, המשמעות של מיטוזה רבה, שכן בזכותה מתאפשרים התפתחות וצמיחה של אורגניזמים חד-תאיים ורב-תאיים, התחדשות של חלקים מסוימים בגוף ורבייה א-מינית.
חידון נושא
הערכת דוח
דירוג ממוצע: 4.6. סך הדירוגים שהתקבלו: 296.
ארגון כללי של מיטוזה
כפי שתיאוריית התאים מניחה, עלייה במספר התאים מתרחשת אך ורק בשל חלוקת התא המקורי, שהכפיל בעבר את החומר הגנטי שלו. זהו האירוע המרכזי בחיי התא כשלעצמו, כלומר השלמת השעתוק מסוגו. כל חיי ה"בין-פאזיים" של תאים מכוונים למימוש מלא של מחזור התא, המסתיים בחלוקת התא. חלוקת התא עצמה היא תהליך לא אקראי, שנקבע גנטית בהחלט, שבו שרשרת שלמה של אירועים מסודרת בשורה ברצף.
כפי שכבר הוזכר, החלוקה של תאים פרוקריוטים ממשיכה ללא עיבוי כרומוזומים, אם כי צריך להיות מספר תהליכים מטבולייםוקודם כל, סינתזות של הסדרה חלבונים ספציפייםמעורב בחלוקה ה"פשוטה" של תא החיידק לשניים.
חלוקה של כולם תאים איקריוטייםעקב עיבוי של כרומוזומים כפולים (משוכפלים), אשר לובשים צורה של מבנים חוטים צפופים. כרומוזומים חוטיים אלו נישאים לתאי בת באמצעות מבנה מיוחד - ציר חלוקה.סוג זה של חלוקת תאים אוקריוטיים הוא מיטוזה(מיוונית. מיטוס- חוטים), או מיטוזה,אוֹ חלוקה עקיפה- היא הדרך השלמה היחידה להגדיל את מספר התאים. חלוקת תאים ישירה, או אמיטוזיס, מתוארת בצורה מהימנה רק בחלוקת מקרו-גרעיניים פוליפלואידים של ריצות, המיקרו-גרעינים שלהם מתחלקים רק על ידי מיטוזה.
החלוקה של כל התאים האיקריוטים קשורה להיווצרות של מיוחד מַנגָנוֹן חלוקת תא. כאשר תאים משתכפלים, מתרחשים שני אירועים: התבדלות של כרומוזומים משוכפלים וחלוקת גוף התא - ציטוטומיה.החלק הראשון של האירוע באוקריוטים מתבצע בעזרת מה שנקרא ציר חלוקה,מורכב ממיקרוטובולים, והחלק השני מתרחש עקב השתתפותם של מתחמי אקטומיוזין, גורם לחינוךהתכווצויות בתאים ממקור בעלי חיים או עקב השתתפותם של מיקרוטובולים וחוטי אקטין ביצירת פרגמופלסט, דופן התא הראשונית בתאי הצמח.
שני סוגים של מבנים לוקחים חלק ביצירת ציר החלוקה בכל התאים האוקריוטיים: גופים קוטביים (קטבים) של הציר וקינטוכורים של כרומוזומים. גופים קוטביים, או צנטרוזומים, הם מרכזי הארגון (או הגרעין) של מיקרוטובולים. מיקרו-צינוריות צומחות מהן עם הקצוות הפלוסים שלהן, ויוצרות צרורות הנמתחות אל הכרומוזומים. בתאי בעלי חיים, הצנטרוזומים כוללים גם צנטרולים. אבל לאאוקריוטים רבים אין צנטריולים, ומרכזי ארגון מיקרו-צינוריות קיימים בצורה של אזורים אמורפיים חסרי מבנה, שמהם משתרעים מיקרו-טובוליות רבות. ככלל, שני צנטרוזומים או שני גופים קוטביים מעורבים בארגון מנגנון החלוקה, הממוקם בקצוות מנוגדים של גוף מורכב בצורת ציר המורכב ממיקרוטובולים. המבנה השני האופייני לחלוקת תאים מיטוטיים, המחבר את מיקרו-צינוריות הציר עם הכרומוזום, הוא kinetochores.זה kinetochores, באינטראקציה עם microtubules, אשר אחראים על תנועת הכרומוזומים במהלך חלוקת התא.
כל המרכיבים הללו, כלומר גופים קוטביים (צנטרוזומים), מיקרו-צינוריות ציר וקינטוכורים כרומוזומליים, נמצאים בכל התאים האוקריוטיים משמרים ועד יונקים, ומספקים תהליך מורכב של הפרדה של כרומוזומים משוכפלים.
סוגים שוניםמיטוזיס איקריוטים
החלוקה של תאי בעלי חיים וצמחים שתוארה לעיל אינה הצורה היחידה של חלוקת תאים עקיפה (איור 299). הסוג הפשוט ביותר של מיטוזה הוא pleuromitosis.במידה מסוימת, הוא דומה לביקוע בינארי של תאים פרוקריוטים, שבו הנוקלואידים לאחר השכפול נשארים קשורים לממברנת הפלזמה, שמתחילה לצמוח בין נקודות הקישור של ה-DNA, ובכך, כביכול, מפזרת את הכרומוזומים לתוך אזורים שוניםתאים (על חלוקת פרוקריוטות, ראה להלן). לאחר מכן, במהלך היווצרות התכווצות התא, כל אחת ממולקולות ה-DNA תהיה בתא נפרד חדש.
כפי שכבר הוזכר, יצירת ציר הבנוי ממיקרוטובוליות אופיינית לחלוקת תאים אוקריוטיים (איור 300). בְּ pleuromitosis סגור(זה נקרא סגור מכיוון שההתבדלות של הכרומוזומים מתרחשת מבלי לשבור את הממברנה הגרעינית) כמרכזי ארגון המיקרו-צינוריות (MCMT), לא צנטריולים משתתפים, אלא מבנים אחרים הממוקמים על בְּתוֹךממברנה גרעינית. אלה הם מה שנקרא גופים קוטביים של מורפולוגיה בלתי מוגדרת, שמהם משתרעים מיקרוטובולים. ישנם שניים מהגופים הללו, הם מתפצלים זה מזה מבלי לאבד את הקשר שלהם עם המעטפת הגרעינית, וכתוצאה מכך נוצרים שני חצי צירים הקשורים לכרומוזומים. כל תהליך היווצרות המנגנון המיטוטי וההתבדלות של הכרומוזומים מתרחשים במקרה זה מתחת לממברנה הגרעינית. סוג זה של מיטוזה נמצא בקרב פרוטוזואה, הוא נפוץ בפטריות (chytridia, zygomycetes, שמרים, oomycetes, ascomycetes, myxomycetes, וכו'). ישנן צורות של pleuromitosis סגורה למחצה, כאשר המעטפת הגרעינית נהרסת בקטבים של הציר שנוצר.
צורה נוספת של מיטוזה היא אורתומיטוזיס. בְּבמקרה זה, ה-COMTs ממוקמים בציטופלזמה; מההתחלה, לא נוצרים סמי-צירים, אלא ציר דו-קוטבי. ישנן שלוש צורות של אורתומיטוזיס: לִפְתוֹחַ(מיטוזה רגילה), חצי סגורו סָגוּר.באורתומיטוזיס סגורה למחצה, נוצר ציר ביסימטרי בעזרת TsOMT הממוקם בציטופלזמה, המעטפת הגרעינית נשמרת לאורך כל המיטוזה, למעט אזורי הקוטב. ניתן למצוא כאן המוני חומר גרגירי או אפילו צנטריולים בשם COMT. צורה זו של מיטוזה מצויה ב-zoospores של אצות ירוקות, חומות ואדומות, בחלק מהפטריות הנמוכות יותר ובגרגרינים. עם אורתומיטוזיס סגור, הממברנה הגרעינית נשמרת לחלוטין, שמתחתיה נוצר ציר אמיתי. מיקרוטובולים נוצרים בקאריופלזמה, לעתים רחוקות יותר הם גדלים מ-COMT תוך גרעיני, שאינו קשור (בניגוד לפלאורומיטוזיס) עם הממברנה הגרעינית. סוג זה של מיטוזה מאפיין את חלוקתם של מיקרוגרעינים ריסים, אך נמצא גם בפרוטוזואים אחרים. באורתומיטוזיס פתוח, המעטפת הגרעינית מתפרקת לחלוטין. סוג זה של חלוקת תאים אופייני לאורגניזמים של בעלי חיים, כמה פרוטוזואה ותאים של צמחים גבוהים יותר. צורה זו של מיטוזה, בתורה, מיוצגת על ידי טיפוסים אסטרליים ואנסטרליים (איור 301).
מסקירה קצרה זו, ברור כי תכונה עיקריתמיטוזיס באופן כללי היא הופעת מבנים של ציר הביקוע, שנוצר בקשר עם TsOMT, שהוא מגוון במבנה.
מורפולוגיה של הדמות המיטוטית
כפי שכבר הוזכר, המנגנון המיטוטי נחקר באופן יסודי ביותר בתאים של צמחים ובעלי חיים גבוהים יותר. זה בא לידי ביטוי טוב במיוחד בשלב המטאפאזה של המיטוזה (ראה איור 300). בתאים חיים או קבועים במטאפאזה, במישור המשווני של התא, ממוקמים כרומוזומים, שמהם מה שנקרא חוטי ציר,מתכנס בשני קטבים שונים של הדמות המיטוטית. אז הציר המיטוטי הוא אוסף של כרומוזומים, קטבים וסיבים. סיבי הציר הם מיקרוטובולים בודדים או צרורותיהם. מיקרו-צינוריות מתחילות מקטבי הציר, וחלקן מגיעות לצנטרומרים, שם ממוקמים הקינטוכורים של הכרומוזומים (kinetochore microtubules), חלקם הולכים יותר לכיוון הקוטב הנגדי, אך אינם מגיעים אליו - "מיקרוטובולים בין-קוטביים". בנוסף, קבוצה של מיקרוטובולים רדיאליים יוצאת מהקטבים, ויוצרת סביבם, כביכול, "זוהר קורן" - אלה הם מיקרוטובולים אסטרליים.
על פי המורפולוגיה הכללית, דמויות מיטוטיות מחולקות לשני סוגים: אסטרלי ואנסטרלי (ראה איור 301).
סוג הציר האסטרלי (או מתכנס) מאופיין בכך שהקטבים שלו מיוצגים על ידי אזור קטן שאליו מתכנסות (מתכנסות) המיקרוטובולים. בדרך כלל, צנטרוזומים המכילים צנטרולים ממוקמים בקטבים של הצירים האסטרליים. אמנם ידועים מקרים של מיטוזות אסטרליות צנטרולריות (במהלך מיוזה של חלק מחסרי חוליות). בנוסף, מיקרוטובולים רדיאליים מתפצלים מהקטבים, שאינם חלק מהציר, אלא יוצרים אזורי כוכבים - ציסטרים. באופן כללי, סוג זה של ציר מיטוטי דומה יותר למשקולת (ראה איור 301, א).
לסוג האנסטריאלי של הדמות המיטוטית אין ציסטרים בקטבים. האזורים הקוטביים של הציר כאן רחבים, הם נקראים כובעים קוטביים, הם לא כוללים centrioles. הסיבים של הציר מקרה זהלא יוצאים מנקודה אחת, אלא מתפצלים בחזית רחבה (מתפצלים) מכל אזור כובעי הקוטב. סוג זה של ציר מאפיין תאים מתחלקים של צמחים גבוהים יותר, אם כי הוא נמצא לעתים בבעלי חיים גבוהים יותר. לפיכך, בעוברות מוקדמת של יונקים, מיטוזות צנטרולריות (מתפצלות) נצפו במהלך חלוקת הבשלת ביציות ובמהלך חלוקות I ו-II של הזיגוטה. אבל החל מחלוקת התא השלישית ובכל אלה שלאחר מכן, תאים מתחלקים בהשתתפות צירים אסטרליים, שבקטבים שלהם נמצאים תמיד צנטרולים.
באופן כללי, עבור כל צורות המיטוזה, הכרומוזומים עם הקינטוכורים שלהם, הגופים הקוטביים (צנטרוזומים) וסיבי הציר נשארים מבנים נפוצים.
צנטרומרים וקינטוכורים
צנטרומרים כאתרי קישור לכרומוזומים עם מיקרוטובולים יכולים להיות לוקליזציה שונהלאורך הכרומוזומים. לדוגמה, הולוצנטריצנטרומרים מתרחשים כאשר מיקרו-צינוריות קשורות לאורך כל הכרומוזום (חלק מהחרקים, נמטודות, חלק מהצמחים) ו מונוצנטרי centromeres - כאשר מיקרוטובולים קשורים לכרומוזומים באזור אחד (איור 302). צנטרומרים מונוצנטריים יכולים להיות לְאַתֵר בִּמְדוּיָק(לדוגמה, בשמרים ניצנים מסוימים), כאשר רק מיקרו-צינורית אחת מתקרבת לקינטוכור, ו אזורי, שבו צרור של מיקרוטובולים מתקרב לקינטוכור המורכב. למרות מגוון אזורי הצנטרומרים, כולם קשורים למבנה מורכב. kinetochore,שיש לו דמיון מהותי במבנה ובתפקוד בכל האיקריוטים.
|
אורז. 302. קינטוכורים באזור הצנטרומרי של הכרומוזומים 1 - kinetochore; 2 - צרור של microtubules kinetochore; 3 - כרומטיד |

המבנה הפשוט ביותר של הקינטוכור המונוצנטרי הוא בתאי שמרי האופה ( Saccharomyces cerevisiae). זה קשור לקטע מיוחד של DNA על הכרומוזום (מוקד צנטרומרי או CEN). אזור זה מורכב משלושה יסודות DNA: CDE I, CDE II, CDE III. מעניין לציין שרצפי הנוקלאוטידים ב-CDE I ו-CDE III שמורים מאוד ודומים לאלה שב-Drosophila. אזור CDE II יכול להיות בגדלים שונים והוא מועשר בזוגות A-T. לקשר עם מיקרוטובולים S. cerevisiaהאתר CDE III, המקיים אינטראקציה עם מספר חלבונים, אחראי.
צנטרומרים אזוריים מורכבים ממוקדי CEN חוזרים ומועשרים באזורים של הטרוכרומטין מכונן המכילים DNA לווייני הקשור לקינטוכורים.
קינטוכורים הם מבני חלבון מיוחדים, הממוקמים ברובם באזורי הצנטרומרים של הכרומוזומים (ראה איור 302). Kinetochores נחקר טוב יותר באורגניזמים גבוהים יותר. Kinetochores הם קומפלקסים מורכבים המורכבים מחלבונים רבים. מבחינה מורפולוגית, הם דומים מאוד, בעלי אותו מבנה, החל מדיאטומים לבני אדם. קינטוכורים הם מבנים תלת-שכבתיים (איור 303): השכבה הפנימית הצפופה הצמודה לגוף הכרומוזום, השכבה הרפויה האמצעית והשכבה הצפופה החיצונית. שריבים רבים משתרעים מהשכבה החיצונית, ויוצרים את מה שנקרא הכתר הסיבי של הקינטוכור (איור 304).
בצורה הכללית, קינטוכורים נראים כמו לוחות או דיסקים השוכבים באזור ההתכווצות הראשונית של הכרומוזום, בצנטרומר. בדרך כלל יש קינטוכור אחד לכל כרומטיד (כרומוזום). לפני אנפאזה, הקינטוכורים על כל כרומטיד אחות מסודרים הפוך, כל אחד מתחבר עם צרור מיקרו-צינוריות משלו. בצמחים מסוימים, הקינטוכור אינו נראה כמו לוחות, אלא כחצי כדור.
Kinetochores הם קומפלקסים מורכבים, שבהם, בנוסף ל-DNA ספציפי, מעורבים חלבוני קינטוכור רבים (CENP proteins) (איור 305). באזור הצנטרומר של הכרומוזום, מתחת לקינטוכור התלת-שכבתי, ישנו אזור של הטרוכרומטין המועשר ב-DNA α-לווין. עוד נמצאים כאן מספר חלבונים: CENP-B, הנקשר ל-α-DNA; MSAC, חלבון דמוי קינזין; כמו גם חלבונים האחראים על הזיווג של כרומוזומי אחיות (קוהזינים). החלבונים הבאים זוהו בשכבה הפנימית של הקינטוכור: CENP-A, וריאנט של ההיסטון H3, הנקשר כנראה לאזור ה-DNA CDE II; CENP-G, הנקשר לחלבוני המטריצה הגרעינית; חלבון CENP-C משומר עם תפקיד לא ידוע. החלבון 3F3/2 נמצא בשכבה הרופפת האמצעית, אשר, ככל הנראה, רושם איכשהו את המתח של צרורות מיקרוטובוליות. בשכבה הצפופה החיצונית של הקינטוכור, זוהו חלבוני CENP-E ו-CENP-F המעורבים בקישור מיקרוטובולי. בנוסף, ישנם חלבונים ממשפחת הדיניינים הציטופלזמיים.
תפקיד פונקציונליקינטוכורים מורכבים בקשירה של כרומטידות אחיות זו לזו, בקיבוע של מיקרוטובוליות מיטוטיות, בוויסות הפרדת הכרומוזומים ובתנועה בפועל של כרומוזומים בזמן מיטוזה בהשתתפות מיקרוטובולים.
מיקרוטובולים הגדלים מהקטבים, מהצנטרוזומים, מתקרבים לקינטוכורים. המספר המינימלי בשמרים - מיקרוטובולה אחת לכל כרומוזום. בצמחים גבוהים יותר, מספר זה מגיע ל-20-40. בְּ בתקופה האחרונההצליח להראות שהקינטוכורים המורכבים של אורגניזמים גבוהים יותר הם מבנה המורכב מתת-יחידות חוזרות, שכל אחת מהן מסוגלת ליצור קשרים עם מיקרוטובולים (איור 306). על פי אחד המודלים של מבנה האזור הצנטרומרי של הכרומוזום (Zinkowski, Meine, Brinkley, 1991), הוצע כי תת-יחידות קינטוכור המכילות את כל החלבונים האופייניים ממוקמות ב-interphase על אזורי DNA ספציפיים. כאשר הכרומוזומים מתעבים בפרופאזה, תת-יחידות אלו מתקבצות בצורה כזו שנוצרת אזור מועשר בקומפלקסים של חלבון אלו, - kinetochore.
קינטוכורים, מבני חלבון באופן כללי, כפולים בתקופת S, במקביל להכפלת הכרומוזומים. אבל החלבונים שלהם נמצאים על הכרומוזומים בכל תקופות מחזור התא (ראה איור 303).
דינמיקה של מיטוזיס
בחלקים רבים של ספר זה, כבר נגענו בהתנהגותם של מרכיבים תאיים שונים (כרומוזומים, נוקלאולי, מעטפת גרעינית וכו') במהלך חלוקת התא. אבל בואו נחזור לאלה. תהליכים קריטייםלהבין אותם כמכלול.
בתאים שנכנסו למחזור החלוקה, שלב המיטוזה עצמו, החלוקה העקיפה, לוקח זמן קצר יחסית, רק כ-0.1 מזמן מחזור התא. אז, בתאים מתחלקים של מריסטם השורש, האינטרפאזה יכולה להיות 16-30 שעות, והמיטוזה יכולה להימשך רק 1-3 שעות. תאי האפיתלמעי העכבר נמשך כ-20-22 שעות, בעוד שהמיטוזה אורכת שעה אחת בלבד.כאשר הביצים נמעכות, כל תקופת התא, כולל מיטוזה, יכולה להיות פחות משעה.
תהליך חלוקת התאים המיטוטי מחולק בדרך כלל למספר שלבים עיקריים: פרופאזה, פרומטפאזה, מטאפאזה, אנפאזה, טלופזה (איור 307-312). קשה מאוד לקבוע את הגבולות בין השלבים הללו במדויק, כי המיטוזה עצמה היא תהליך מתמשך ושינוי השלבים מתרחש בהדרגה רבה: אחד מהם עובר באופן בלתי מורגש לאחר. השלב היחיד שיש לו התחלה אמיתית הוא אנפאזה - תחילת תנועת הכרומוזומים לכיוון הקטבים. משך השלבים האישיים של המיטוזה שונה, הקצר ביותר בזמן הוא האנפאזה (טבלה 15).
הזמן של שלבים בודדים של מיטוזה נקבע בצורה הטובה ביותר על ידי התבוננות ישירה של חלוקת תאים חיים בחדרים מיוחדים. לדעת את זמן המיטוזה, אפשר לחשב את משך שלבים בודדים לפי אחוז התרחשותם בין תאים מתחלקים.
פרופאזה.כבר בסוף תקופת G 2 מתחילים להתרחש סידורים משמעותיים מחדש בתא. אי אפשר לקבוע בדיוק מתי מתרחשת הפרופזה. הקריטריון הטוב ביותר לתחילת שלב זה של מיטוזה יכול להיות ההופעה בגרעינים של מבנים חוטים - כרומוזומים מיטוטיים. לאירוע זה מקדימה עלייה בפעילות של פוספורילאזים המשנים היסטונים, בעיקר היסטון H1. בפרופאזה, כרומטידות אחיות מקושרות זו לצד זו בעזרת חלבוני קוהזין, היוצרים את הקשרים הללו כבר בתקופת S, במהלך שכפול הכרומוזומים. לפי הנבואה המאוחרת, הקשר בין כרומטידות אחיות נשמר רק באזור הקינטוכורים. בכרומוזומי פרופאזה כבר ניתן לראות קינטוכורים בוגרים, שאין להם קשרים עם מיקרוטובולים.
עיבוי הכרומוזומים בגרעין הפרופזה חופף לירידה חדה בפעילות התעתיק של הכרומטין, שנעלם לחלוטין באמצע הפרופזה. עקב ירידה בסינתזה של RNA ועיבוי כרומטין, מתרחשת גם נטרול של גנים גרעיניים. במקביל, מרכזי פיברילרים בודדים מתמזגים בצורה כזו שהם הופכים למקטעים יוצרים גרעיניים של כרומוזומים, למארגנים גרעיניים. רוב החלבונים הגרעיניים מתנתקים ונמצאים בצורה חופשית בציטופלזמה של התא או נקשרים לפני השטח של הכרומוזומים.
במקביל, מתרחשת זרחון של מספר חלבוני lamina - המעטפת הגרעינית, שמתפרקת. במקרה זה אבד הקשר של המעטפת הגרעינית עם הכרומוזומים. ואז המעטפת הגרעינית מפוצלת ל-vacuoles קטנים, ומתחמי הנקבוביות נעלמים.
במקביל לתהליכים אלו, נצפית הפעלה של מרכזי תאים. בתחילת הפרופזה, מפורקים מיקרוטובולים בציטופלזמה ו צמיחה נפיצהמיקרוטובולים אסטרליים רבים סביב כל אחד מהדיפלוזומים הכפולים (איור 308). קצב הגדילה של מיקרוטובוליות בפרופאזה גבוה כמעט פי שניים מהצמיחה של מיקרוטובוליות בין-פאזיות, אך רגישותן גבוהה פי 5-10 מזו של ציטופלזמה. אז אם זמן מחצית החיים של המיקרוטובולים בציטופלזמה הוא בערך 5 דקות, אז במהלך המחצית הראשונה של המיטוזה הוא רק 15 שניות. כאן, חוסר היציבות הדינמית של המיקרוטובולים בולטת עוד יותר. כל המיקרוטובולים המשתרעים מהצנטרוזומים צומחים קדימה עם קצוות הפלוס שלהם.
צנטרוזומים פעילים - עמודי הציר העתידיים - מתחילים להתפצל זה מזה למרחק מסוים. המנגנון של התבדלות פרופאזה כזו של הקטבים הוא כדלקמן: מיקרוטובולים אנטי-מקבילים הנעים זה לכיוון השני מקיימים אינטראקציה זה עם זה, מה שמוביל לייצוב ולדחייה גדולים יותר של הקטבים (איור 313). זה מתרחש עקב האינטראקציה עם מיקרו-צינוריות של חלבונים דמויי דיניין, אשר בחלק המרכזי של הציר מסדרים מיקרו-צינוריות בין-קוטביות במקביל זו לזו. במקביל נמשכים הפילמור וצמיחתם, המלווים בדחיפתם לכיוון הקטבים עקב עבודתם של חלבונים דמויי קינזין (איור 314). בשלב זה, במהלך היווצרות הציר, המיקרוטובולים עדיין אינם קשורים לקינטוכורים של הכרומוזומים.
בפרופאזה, במקביל לפירוק המיקרוטובולים הציטופלזמיים, הרטיקולום האנדופלזמי אינו מאורגן (הוא מתפרק לוואקווולים קטנים השוכנים לאורך פריפריית התא) ומנגנון גולגי, המאבד את הלוקליזציה הפרי-גרעינית שלו, מחולק לדיקטיוזומים נפרדים המפוזרים באקראי בציטופלזמה. .
פרומטפאזה.לאחר הרס המעטפת הגרעינית, הכרומוזומים המיטוטים שוכנים באזור הגרעין הקודם ללא סדר מסוים. בפרומטפאזה מתחילה תנועתם ותנועתם, מה שמוביל בסופו של דבר ליצירת "לוחית" כרומוזום משווני, לסידור מסודר של כרומוזומים בחלק המרכזי של הציר כבר במטאפאזה. בפרומטפאזה, ישנה תנועה מתמדת של כרומוזומים, או מטאקינזיס, שבה הם מתקרבים אל הקטבים, או משאירים אותם לכיוון מרכז הציר עד שהם תופסים את המיקום האמצעי האופייני למטאפאזה (קונגרס הכרומוזומים).
בתחילת הפרומטאפאזה, הכרומוזומים השוכנים קרוב יותר לאחד הקטבים של הציר שנוצר מתחילים להתקרב אליו במהירות. זה לא קורה בבת אחת, אלא לוקח פרק זמן מסוים. נמצא שסחיפה אסינכרונית ראשונית כזו של כרומוזומים לקטבים שונים מתבצעת בעזרת מיקרוטובולים. באמצעות הגברה ניגודיות פאזה וידאו-אלקטרונית במיקרוסקופ אור, ניתן היה לראות על תאים חיים שמיקרוטובולים בודדים המשתרעים מהקטבים מגיעים בטעות לאחד מהקינטוכורים של הכרומוזום ונקשרים אליו, "נלכדים" על ידי הקינטוכור. לאחר מכן, גלישת כרומוזומים מהירה, בקצב של כ-25 מיקרומטר לדקה, לאורך המיקרוטובול לכיוון הקצה המינוס שלה. זה מוביל לעובדה שהכרומוזום מתקרב לקוטב שממנו הגיעה המיקרוטובולה הזו (איור 315). חשוב לציין כי kinetochores יכולים ליצור קשר עם פני השטח לרוחב של microtubules כאלה. במהלך תנועה זו של הכרומוזומים, המיקרוטובולים אינם מפורקים. סביר להניח שחלבון מוטורי הדומה לדינאין הציטופלזמי המצוי בכתר הקינטוכורים אחראי לתנועה כה מהירה של כרומוזומים.
כתוצאה מתנועת פרומטפאזה ראשונית זו, הכרומוזומים מתקרבים באופן אקראי אל קטבי הציר, שם נמשכת היווצרותם של מיקרוטובולים חדשים. ברור שככל שהקינטוכור הכרומוזומלי קרוב יותר לצנטרוזום, כך גדלה האקראיות של האינטראקציה שלו עם מיקרוטובולים אחרים. במקרה זה, קצוות פלוס חדשים וגדלים של מיקרוטובולים "נלכדים" על ידי האזור של כתר הקינטוכור; כעת מחוברת צרור של מיקרו-צינוריות לקינטוכור, שצמיחתן ממשיכה בקצה הפלוס שלהן. עם צמיחתו של צרור כזה, הקינטוכור, ואיתו הכרומוזום, חייבים לנוע לכיוון מרכז הציר, להתרחק מהקוטב. אבל בשלב זה, מיקרוטובולים צומחים מהקוטב הנגדי לקינטוכור השני של הכרומטיד האחות השני, שהצרור שלו מתחיל למשוך את הכרומוזום לקוטב הנגדי. נוכחותו של כוח משיכה כזה מוכחת על ידי העובדה שאם חותכים צרור של מיקרו-צינוריות באחד מהקינטוכורים באמצעות מיקרו-קרן לייזר, אז הכרומוזום מתחיל לנוע לכיוון הקוטב הנגדי (איור 316). בתנאים רגילים, הכרומוזום, המבצע תנועות קטנות לכיוון הקוטב האחד או השני, כתוצאה מכך תופס בהדרגה מיקום אמצעי בציר. בתהליך של סחף כרומוזומי פרומטפאזה, המיקרוטובוליות מתארכות ונבנות בקצוות הפלוס כאשר הקינטוכור מתרחק מהקוטב, ומיקרוטובוליות מפורקות ומתקצרות גם בקצה הפלוס, כאשר הקינטוכור האחות נעה לכיוון הקוטב. .
תנועות מתחלפות אלו של הכרומוזומים פה ושם מובילות לעובדה שבסופו של דבר הם מגיעים לקו המשווה של הציר ומסתדרים בשורה בלוח המטאפאזה (ראה איור 315).
מטאפאזה(איור 309). במטאפאזה, כמו גם בשלבים אחרים של מיטוזה, למרות ייצוב מסוים של צרורות המיקרוטובוליות, חידושם המתמיד נמשך עקב הרכבה ופירוק של טובולינים. במהלך המטאפאזה, הכרומוזומים מסודרים כך שהקינטוכורים שלהם פונים לקטבים מנוגדים. יחד עם זאת, יש מחיצה קבועה ומיקרוטובולים בין-קוטביים, שמספרם במטאפאזה מגיע למקסימום. אם מסתכלים על תא המטאפאזה מהצד של הקוטב, אז אפשר לראות שהכרומוזומים מסודרים כך שהקטעים הצנטרומריים שלהם פונים למרכז הציר, והכתפיים פונות לפריפריה. סידור זה של כרומוזומים נקרא "כוכב האם" והוא אופייני לתאי בעלי חיים (איור 317). בצמחים, לעתים קרובות במטאפאזה, הכרומוזומים שוכנים במישור המשווני של הציר ללא סדר קפדני.
בסוף המטאפאזה, תהליך ההפרדה של כרומטידות אחיות זו מזו הושלם. הכתפיים שלהם מקבילות זו לזו, הפער המפריד ביניהם נראה בבירור ביניהן. המקום האחרון שבו נשמר המגע בין הכרומטידות הוא הצנטרומר; עד סוף המטאפאזה, הכרומטידות בכל הכרומוזומים נשארות מחוברות באזורים הצנטרומריים.
אנפאזהמתחיל בפתאומיות, שניתן לראות היטב במחקר החיוני. אנפאזה מתחילה בהפרדה של כל הכרומוזומים בבת אחת באזורים הצנטרומריים. בזמן זה, ישנה השפלה בו-זמנית של קוהזינים צנטרומרים, שעד לאותו זמן קשרו כרומטידות אחיות. הפרדה בו-זמנית זו של הכרומטידים מאפשרת להם להתחיל את ההפרדה הסינכרונית שלהם. כרומוזומים מאבדים לפתע את הרצועות הצנטרומריות שלהם ומתחילים להתרחק זה מזה באופן סינכרוני לכיוון הקטבים המנוגדים של הציר (איור 310 ו-318). מהירות תנועת הכרומוזומים אחידה, היא יכולה להגיע ל-0.5-2 מיקרומטר/דקה. אנפאזה היא השלב הקצר ביותר של מיטוזה (כמה אחוזים מהזמן הכולל), אך במהלך תקופה זו מתרחשים מספר אירועים. העיקריים שבהם הם הפרדה של שתי קבוצות זהות של כרומוזומים והובלתם לקצוות מנוגדים של התא.
|
אורז. 318. סטייה אנפאזית של כרומוזומים א - אנפאזה א'; 6 - אנפאזה ב' |

בעת תנועה, הכרומוזומים משנים את הכיוון שלהם ולעתים קרובות מקבלים צורת V. החלק העליון שלהם מופנה לכיוון מוטות החלוקה, והכתפיים נזרקות, כביכול, חזרה למרכז הציר. אם שבר בזרוע הכרומוזומים התרחש לפני אנפאזה, אז במהלך האנפאזה הוא לא ישתתף בתנועת הכרומוזומים ויישאר באזור המרכזי. תצפיות אלו הראו שהאזור הצנטרומרי, יחד עם הקינטוכור, הוא זה שאחראי לתנועת הכרומוזומים. נראה שהכרומוזום נמשך אל הקוטב שמעבר לצנטרומר. בחלק מהצמחים הגבוהים יותר (אוסיקה) אין התכווצות צנטרומרית בולטת, וסיבי הציר נמצאים במגע עם נקודות רבות על פני הכרומוזומים (כרומוזומים פולצנטריים והולוצנטריים). במקרה זה, הכרומוזומים ממוקמים על פני סיבי הציר.
למעשה, ההתבדלות של הכרומוזומים מורכבת משני תהליכים: 1 - התבדלות הכרומוזומים עקב צרורות קינטוכורים של מיקרוטובולים; 2 - התבדרות הכרומוזומים יחד עם הקטבים עקב התארכות המיקרוטובולים הבין-קוטביים. הראשון מבין התהליכים הללו נקרא "אנפאזה A", השני - "אנפאזה B" (ראה איור 318).
במהלך אנפאזה A, כאשר קבוצות של כרומוזומים מתחילות לנוע לכיוון הקטבים, מתרחש קיצור של צרורות הקינטוכור של המיקרוטובולים. ניתן היה לצפות שבמקרה זה דה-פולימריזציה של מיקרוטובולים תתרחש בקצוות המינוס שלהם; מסתיים הכי קרוב למוט. עם זאת, הוכח שמיקרוטובולים אכן מתפרקים, אך בעיקר (80%) מקצוות הפלוס הסמוכים לקינטוכורים. בניסוי, טובולין הקשור ל-fluorochrome הוכנס לתאי תרבית רקמה חיים בשיטת המיקרו-הזרקה. זה איפשר לראות באופן חיוני את המיקרוטובולים בציר הביקוע. בתחילת האנפאזה, צרור הציר של אחד הכרומוזומים הוקרן במיקרו-קרן קלה בערך באמצע בין הקוטב לכרומוזום. עם חשיפה זו, הקרינה נעלמת באזור המוקרן. תצפיות הראו שהאזור המוקרן אינו מתקרב אל הקוטב, אך הכרומוזום מגיע אליו כאשר צרור הקינטוכור מתקצר (איור 319). כתוצאה מכך, פירוק המיקרוטובולים של צרור הקינטוכור מתרחש בעיקר מהקצה הפלוס, בנקודת החיבור שלו עם הקינטוכור, והכרומוזום נע לעבר קצה המינוס של המיקרוטובולים, הנמצא באזור הצנטרוזום. התברר שתנועה כזו של כרומוזומים תלויה בנוכחות ATP ובנוכחות של ריכוז מספיק של יוני Ca 2+. העובדה שחלבון הדיניין נמצא בהרכב הכתר הקינטוכור, שבו משובצים קצוות הפלוסים של המיקרוטובולים, אפשרה לנו להניח שהמנוע הוא שמושך את הכרומוזום אל הקוטב. במקביל לכך, מתרחשת דה-פולימריזציה של microtubules kinetochore בקצה הפלוס (איור 320).
לאחר הפסקת הכרומוזומים בקטבים, נצפית התבדלות נוספת שלהם עקב הוצאת הקטבים זה מזה (אנפאזה B). הוכח שבמקרה זה, הקצוות הפלוסים של המיקרוטובולים הבין-קוטביים גדלים, אשר יכולים להגדיל משמעותית את אורכם. האינטראקציה בין המיקרוטובולים האנטי-מקבילים הללו, המובילה להחלקתן ביחס זו לזו, נקבעת על ידי חלבונים דמויי קינזין מוטוריים אחרים. בנוסף, הקטבים נמשכים בנוסף לפריפריה של התא עקב אינטראקציה עם מיקרוטובולים אסטרליים של חלבונים דמויי דיניין על קרום הפלזמה.
רצף האנפאזות A ו-B ותרומתם לתהליך הפרדת הכרומוזומים יכולים להיות שונים באובייקטים שונים. אז, ביונקים, שלבים A ו-B מתרחשים כמעט בו זמנית. בפרוטוזואה, אנפאזה B יכולה לגרום לעלייה של פי 15 באורך הציר. שלב ב' נעדר בתאי הצמח.
טלופאזמתחיל בעצירת כרומוזומים (טלופאזה מוקדמת, אנפאזה מאוחרת) (איור 311 ו-312) ומסתיים עם תחילת שחזור של גרעין בין-פאזי חדש (תקופת G 1 מוקדמת) וחלוקת התא המקורי לשני תאי בת (ציטוקינזיס ).
בטלופאזה המוקדמת, הכרומוזומים, מבלי לשנות את כיוונם (אזורים צנטרומריים - לכיוון הקוטב, אזורים טלומריים - לכיוון מרכז הציר), מתחילים להתעבות ולהגדיל את נפחם. באתרי המגע שלהם עם שלפוחיות ממברנות של הציטופלזמה, מתחיל להיווצר קרום גרעיני חדש, שנוצר תחילה על המשטחים הצדדיים של הכרומוזומים ובהמשך באזורים הצנטרומריים והטלומריים. לאחר סגירת הממברנה הגרעינית, מתחילה היווצרותם של נוקלאולים חדשים. התא נכנס לתקופת G 1 של שלב ביניים חדש.
בטלופאזה מתחיל ונגמר תהליך ההרס של המנגנון המיטוטי - פירוק המיקרוטובולים. זה עובר מהקטבים לקו המשווה תא לשעבר: זה בחלק האמצעי של הציר שהמיקרוטובולים נמשכים הכי הרבה זמן (שיורי גוף).
אחד האירועים העיקריים של טלופאז הוא חלוקת גוף התא, כלומר. ציטוטומיה,אוֹ ציטוקינזיס.כבר נאמר לעיל כי בצמחים, חלוקת תאים מתרחשת על ידי היווצרות תוך תאית של מחיצת תאים, ובתאים של בעלי חיים, על ידי התכווצות, החדרה של קרום הפלזמה לתוך התא.
מיטוזה לא תמיד מסתיימת בחלוקת גוף התא. לפיכך, באנדוספרם של צמחים רבים, במשך זמן מה, יכולים להתרחש מספר תהליכים של ביקוע גרעיני מיטוטי ללא חלוקת ציטופלזמה: נוצר סימפלסט רב-גרעיני ענק. כמו כן, ללא ציטוטומיה, גרעינים רבים של פלסמודיה של מיקומיציטים מתחלקים באופן סינכרוני. על שלבים מוקדמיםבמהלך התפתחות העוברים של חרקים מסוימים, מתבצע גם ביקוע חוזר של הגרעינים ללא חלוקת הציטופלזמה.
ברוב המקרים, היווצרות ההיצרות במהלך חלוקת תאי בעלי חיים מתרחשת אך ורק במישור המשווני של הציר. כאן, בסוף האנפאזה, בתחילת הטלופזה, נוצרת הצטברות קליפת המוח של מיקרופילמנטים, היוצרים טבעת מתכווצת (ראה איור 258). המיקרופילמנטים של הטבעת כוללים סיבי אקטין ומולקולות קצרות בצורת מוט של מיוזין II מפולמר. החלקה הדדית של רכיבים אלו מביאה לירידה בקוטר הטבעת ולהופעת חריטה של קרום הפלזמה, הגורמת לבסוף להתכווצות התא המקורי לשניים.
לאחר ציטוטומיה, שני תאים חדשים (בת) נכנסים לשלב G 1, תקופת התא. בשלב זה, הסינתזות הציטופלזמיות מתחדשות, המערכת הוואקואולרית משוחזרת, הדיקטיוזומים של מנגנון גולגי מתרכזים שוב באזור הפרי-גרעיני בשיתוף עם הצנטרוזום. מהצנטרוזום מתחילה צמיחתם של מיקרוטובולים ציטופלזמיים ושיקום הציטושלד הבין-פאזי.
ארגון עצמי של מערכת המיקרוטובוליות
סקירה של היווצרות המנגנון המיטוטי מראה כי הרכבה של מכלול מורכב של מיקרוטובוליות דורשת נוכחות של מרכזי ארגון מיקרוטובוליים וכרומוזומים כאחד.
עם זאת, ישנן מספר דוגמאות המראות כי היווצרותם של ציסטרים וצירים יכולה להתקדם באופן עצמאי, באמצעות ארגון עצמי. אם, בעזרת מיקרומניפולטור, מנותקים חלק מהציטופלזמה הפיברובלסטית, שבו הצנטריול לא יהיה ממוקם, אז מתרחש ארגון מחדש ספונטני של מערכת המיקרוטובוליות. בתחילה, בשבר החתוך, הם מסודרים בצורה כאוטי, אך לאחר זמן מה הם מתאספים עם קצותיהם למבנה דמוי כוכב - ציסטר, שבו קצוות הפלוס של המיקרוטובולים ממוקמים בשולי שבר התא (איור 1). 321). תמונה דומה נצפית בשברים לא-צנטריולים של מלנופורים - תאי פיגמנט הנושאים גרגירי פיגמנט מלנין. במקרה זה, לא רק ההרכבה העצמית של הציטאסטר מתרחשת, אלא גם הצמיחה של מיקרוטובולים מגרגרי פיגמנט שנאספו במרכז שבר התא.
במקרים אחרים, הרכבה עצמית של מיקרוטובולים יכולה להוביל להיווצרות צירים מיטוטיים. אז, באחד הניסויים, הציטוזול בודד מהביצים המתחלקות של קסנופוס. אם שמים בתכשיר כזה כדורים קטנים מכוסים בדנ"א פאג'י, אזי נוצרת דמות מיטוטית, שבה את מקומם של הכרומוזומים תופסים כדורי הדנ"א הללו, שאין להם רצפי קינטוכור, ושני חצאי צירים צמודים אליהם, בקטבים. מהם אין COMTs.
דפוסים דומים נצפים בתנאים טבעיים. לדוגמה, במהלך חלוקה של ביצת תסיסנית בהיעדר צנטריולים, המיקרוטובולים מתחילים להתפלמר באופן אקראי סביב קבוצה של כרומוזומי פרומטפאזה, אשר לאחר מכן מתארגנים מחדש לציר דו-קוטבי ונקשרים לקינטוכורים. תמונה דומה מציינת במהלך החלוקה המיוטית של ביצת הקסנופוס. גם כאן מתרחש תחילה התארגנות ספונטנית של מיקרו-צינוריות שאינן מכוונות סביב קבוצת כרומוזומים, ובהמשך נוצר ציר דו-קוטבי רגיל, שגם בקטבים שלו אין צנטרוזומים (איור 322).
תצפיות אלו הובילו למסקנה שחלבונים מוטוריים, דמויי קינזין ודיניין, לוקחים חלק בארגון העצמי של המיקרוטובולים. נמצאו חלבונים מוטוריים פלוס-טרמינליים - כרומוקינסינים,הקושרים את הכרומוזומים למיקרוטובולים וגורמים לאלה לנוע לכיוון קצה המינוס, מה שמוביל להיווצרות מבנה מתכנס כמו קוטב הציר. מצד שני, מנועים דמויי דיניין הקשורים ל-vacuoles או גרגירים יכולים להזיז מיקרו-צינוריות כך שקצוות המינוס שלהם נוטים ליצור צרורות בצורת חרוט, מתכנסים במרכז החצאי-צירים (איור 323). תהליכים דומים מתרחשים במהלך היווצרות צירים מיטוטיים בתאי צמחים.
מיטוזה של תאי צמח
לחלוקת תאים מיטוטיים של צמחים גבוהים יותר יש מספר של תכונות מאפיינותהקשורים להתחלה ולסיום של תהליך זה. בתאים בין-פאזיים של מריסטמים צמחיים שונים, microtubules ממוקמים בשכבת תת-ממברנה קליפת המוח של הציטופלזמה, ויוצרים צרורות טבעת של microtubules (איור 324). מיקרוטובולים היקפיים נמצאים במגע עם אנזימים היוצרים סיבים תאית, עם סינתטאז של תאית, שהם חלבונים אינטגרליים של קרום הפלזמה. הם מסנתזים תאית על פני ממברנת הפלזמה. מאמינים שבמהלך הצמיחה של סיב התאית, אנזימים אלה נעים לאורך המיקרוטובולים התת-ממברניים.
סידור מחדש מיטוטי של אלמנטים ציטו-שלד מתרחש בתחילת הפרופזה. במקביל, מיקרוטובוליות נעלמות בשכבות ההיקפיות של הציטופלזמה, אך מופיעה צרור טבעתי של מיקרוטובולים בשכבה הקרובה לממברנה של הציטופלזמה באזור המשווה של התא - טבעת preprophase,הכולל יותר מ-100 מיקרוטובולים (איור 325). מבחינה אימונוכימית, גם בטבעת זו נמצא אקטין. חשוב לציין שהטבעת הקדם-פרופאזית של המיקרוטובולים ממוקמת במקום בו תיווצר מחיצת תאים בטלופאז המפרידה בין שני תאים חדשים. מאוחר יותר בפרופזה, הטבעת הזו מתחילה להיעלם, ומיקרוטובוליות חדשות מופיעות לאורך הפריפריה של גרעין הפרופזה. מספרם גדול יותר באזורי הקוטב של הגרעינים; הם, כביכול, עוטפים את כל הפריפריה הגרעינית. במהלך המעבר ל-prometaphase, מתעורר ציר דו-קוטבי, שהמיקרוטובולים שלו מתקרבים למה שנקרא כובעים קוטביים, שבהם נצפים רק וואקוולים קטנים וסיבים דקים של מורפולוגיה בלתי מוגדרת; לא נמצאו סימנים של צנטריולים באזורי הקוטב הללו. כך נוצר הציר האנאסטרלי.
בפרומטפאזה, במהלך חלוקת תאי הצמח, נצפית גם סחיפה מורכבת של כרומוזומים, תנודה ותנועה שלהם מאותו סוג המתרחשות בפרומטפאזה של תאי בעלי חיים. אירועים באנפאזה דומים לאלו במיטוזה אסטרלית. לאחר התפצלות הכרומוזומים, נוצרים גרעינים חדשים, גם עקב פירוק הכרומוזומים והיווצרות מעטפת גרעינית חדשה.
תהליך הציטוטומיה של תאים צמחיים שונה באופן חד מחלוקת התכווצות של תאים ממקור בעלי חיים (איור 326). במקרה זה, פירוק של microtubules ציר באזורי הקוטב מתרחש גם בסוף הטלופזה. אבל המיקרו-צינוריות של החלק העיקרי של הציר בין שני הגרעינים החדשים נשארות, יתר על כן, נוצרות כאן מיקרו-צינוריות חדשות. כך נוצרות צרורות של מיקרו-צינוריות, שאליהן משוייכים ואקוולים קטנים רבים. ואקוולים אלו מקורם בוואקואולים של מנגנון גולגי ומכילים חומרים פקטין. בעזרת מיקרו-צינוריות עוברים וואקוולים רבים לאזור המשווה של התא, שם הם מתמזגים זה עם זה ויוצרים ואקוול שטוח באמצע התא - פרגמופלסט שצומח לכיוון פריפרית התא, כולל עוד ועוד. vacuoles (איור 324, 325 ו-327).
כך נוצר דופן התא הראשוני. בסופו של דבר, קרומי הפרגמופלסט מתמזגים עם קרום הפלזמה: שני תאים חדשים נפרדים, מופרדים על ידי דופן תא חדש שנוצר. ככל שהפרגמופלסט מתרחב, צרורות המיקרוטובוליות נעות יותר ויותר לעבר פריפריית התא. סביר להניח שתהליך מתיחה של הפרגמופלסט והעברת צרורות המיקרוטובולים לפריפריה מקל על ידי צרורות של חוטי אקטין הנמשכים מהשכבה הקורטיקלית של הציטופלזמה במקום שבו הייתה טבעת הקדם-פרופאזה.
לאחר חלוקת התא, המיקרוטובוליות המעורבות בהובלת ואקוולים קטנים נעלמים. דור חדש של microtubules interphase נוצר על הפריפריה של הגרעין, ולאחר מכן ממוקם בשכבת הממברנה הקורטיקלית של הציטופלזמה.
טקובו תיאור כלליחלוקה של תאי צמחים, אך תהליך זה אינו מובן בצורה גרועה ביותר. באזורי הקוטב של הצירים לא נמצאו חלבונים שהם חלק מה-COMT של תאי בעלי חיים. נמצא שבתאי צמחים ניתן למלא תפקיד זה על ידי הממברנה הגרעינית, שממנה מופנים קצוות הפלוס של המיקרוטובולים לפריפריה של התא, וקצוות המינוס אל הממברנה הגרעינית. כאשר הציר נוצר, צרורות הקינטוכור מכוונים עם קצה המינוס אל המוט, וקצה הפלוס אל הקינטוכורים. כיצד מתרחש כיוון מחדש זה של מיקרוטובולים עדיין לא ברור.
במהלך המעבר לפרופזה, מופיעה מסביב לגרעין רשת צפופה של מיקרו-צינוריות, הדומה לסל, אשר לאחר מכן מתחילה להידמות לציר בצורתו. במקרה זה, מיקרוטובולים יוצרים סדרה של צרורות מתכנסים המכוונים לקטבים. מאוחר יותר בפרומטאפאזה מתרחש הקשר של מיקרוטובולים עם קינטוכורים. במטאפאזה יכולים להיווצר סיבים של קינטוכור מרכז משותףהתכנסות - מיני קוטבים של הציר, או מרכזי התכנסות של מיקרוטובולים. סביר להניח, היווצרות של minipoles כאלה מתבצעת על ידי שילוב של הקצוות המינוס של microtubules הקשורים kinetochores. ככל הנראה, בתאים של צמחים גבוהים יותר, תהליך הארגון מחדש של השלד הציטוטי, כולל היווצרות הציר המיטוטי, קשור לארגון העצמי של המיקרוטובולים, אשר, כמו בתאי בעלי חיים, מתרחש בהשתתפות חלבונים מוטוריים.
תנועה וחלוקה של תאי חיידקים
חיידקים רבים מסוגלים לתנועה מהירה בעזרת דגמי חיידקים מיוחדים, או דגלים. צורת התנועה העיקרית של חיידקים היא בעזרת דגלון. הדגלים של חיידקים שונים מהותית מהדגלים של תאים אוקריוטיים. לפי מספר הדגלים, הם מחולקים ל: מונוטריכיים - עם דגלים אחדים, פוליטריים - עם צרור דגלים, פריטריכיים - עם דגלים רבים בחלקים שונים של פני השטח (איור 328).
לפגלות חיידקיות מבנה מורכב מאוד; הם מורכבים משלושה חלקים עיקריים: חוט חיצוני ארוך גלי (הדגל עצמו), קרס וגוף בסיסי (איור 329).
חוט הדגל בנוי מהחלבון flagellin. משקלו המולקולרי משתנה בהתאם לסוג החיידקים (40-60 אלף). תת-היחידות הכדוריות של הפלגלין מתפלמרות לחוטים מעוותים בצורה סלילנית, כך שנוצר מבנה צינורי (לא להתבלבל עם מיקרו-צינוריות אוקריוטיות!) בקוטר של 12-25 ננומטר, חלול מבפנים. הפלגלינים אינם מסוגלים לזוז. הם יכולים להתפלמר באופן ספונטני לחוטים עם גובה גל קבוע האופייני לכל מין. בתאי חיידקים חיים, דגלים גדלים בקצה הרחוק שלהם; כנראה, דגלים מועברים דרך האמצע החלול של הדגלון.
סמוך לפני השטח של התא עובר חוט הפלגלה, הפלגלה, לאזור רחב יותר, מה שנקרא וו. אורכו כ-45 ננומטר והוא מורכב מחלבון אחר.
לגוף הבסיסי החיידקי אין שום קשר לגוף הבסיסי של התא האוקריוטי (ראה איור 290, ב, ג). הוא מורכב ממוט המחובר לוו וארבע טבעות - דיסקים. שתי הטבעות העליונות של הדיסק המצויות בחיידקים גרם שליליים ממוקמות בדופן התא: טבעת אחת (L) טבולה בקרום הליפוסכריד, והשנייה (P) בשכבת ה-murein. שתי הטבעות האחרות, קומפלקס החלבונים S-stator ו-M-רוטור, ממוקמות בממברנת הפלזמה. סמוך לקומפלקס זה בצד קרום הפלזמה שורה עגולה של חלבוני Mot A ו-B.
בגופים הבסיסיים של חיידקים גראם חיוביים, יש רק שתי טבעות תחתונות הקשורות לממברנת הפלזמה. ניתן להבחין בין גופים בזאליים יחד עם ווים. התברר שהם מכילים כ-12 חלבונים שונים.
עקרון התנועה של דגלים חיידקיים שונה לחלוטין מזה של אוקריוטים. אם באאוקריוטים הדגלים נעים עקב החלקה אורכית של כפילות מיקרוטובוליות, אזי בחיידקים תנועת הדגלים מתרחשת עקב סיבוב הגוף הבסיסי (כלומר, דיסקים S ו-M) סביב צירו במישור של קרום פלזמה.
זה הוכח על ידי מספר ניסויים. לכן, על ידי קיבוע הפלגלה על המצע באמצעות נוגדנים לפלגלין, החוקרים צפו בסיבוב של חיידקים. צוין כי מוטציות רבות בפלגלינים (שינויים בכיפוף החוט, "סלסול" וכו') אינן משפיעות על יכולת התנועה של התאים. מוטציות בחלבונים של הקומפלקס הבסיסי מובילות לרוב לאובדן תנועה.
התנועה של דגלים חיידקיים אינה תלויה ב-ATP, אלא מתבצעת עקב שיפוע הטרנסממברני של יוני מימן על פני קרום הפלזמה. במקרה זה, הדיסק M מסתובב.
בסביבת הדיסק M, חלבוני Mot מסוגלים להעביר יוני מימן מהחלל הפריפלסמי לציטופלזמה (עד 1000 יוני מימן מועברים בסיבוב אחד). במקרה זה, הדגלים מסתובבים במהירות אדירה - 5-100 סל"ד, מה שמאפשר לתא החיידק לנוע במהירות 25-100 מיקרון/שניה.
בדרך כלל, חלוקת תאי חיידקים מתוארת כ"בינארית": לאחר שכפול, הנוקלואידים הקשורים לממברנת הפלזמה מתפצלים עקב מתיחה של הממברנה בין הנוקלואידים, ואז נוצרת כיווץ, או מחיצות, המחלקות את התא לשניים. סוג זה של חלוקה מביא לפיזור מדויק מאוד של החומר הגנטי, ללא שגיאות למעשה (פחות מ-0.03% תאים פגומים). נזכיר שהמנגנון הגרעיני של חיידקים, הנוקלואיד, הוא מולקולת DNA ענקית מחזורית (1.6 מ"מ) שיוצרת תחומי לולאה רבים במצב של סליל-על; סדר הערימה של תחומי לולאה אינו ידוע.
הזמן הממוצע בין חלוקה של תאי חיידקים הוא 20-30 דקות. במהלך תקופה זו אמורים להתרחש מספר אירועים: שכפול של ה-DNA הנוקלואיד, הפרדה, הפרדה של נוקלואידים אחים, התבדלות נוספת שלהם, ציטוטומיה עקב היווצרות מחיצה המחלקת את התא המקורי בדיוק לשניים.
כל התהליכים הללו נחקרו באינטנסיביות בשנים האחרונות, ובשל כך התקבלו תצפיות חשובות ובלתי צפויות. אז, התברר שבתחילת סינתזת ה-DNA, שמתחילה מנקודת השכפול (המקור), שתי מולקולות ה-DNA הגדלות נשארות בתחילה קשורות לממברנת הפלזמה (איור 330). במקביל לסינתזת ה-DNA, תהליך הסרת ה-Supercoiling של תחומי הלולאה הישנים והמשכפלים מתרחש עקב מספר אנזימים (טופואיזומראז, גיראזה, ליגאז וכו'), מה שמוביל לבידוד פיזי של שני כרומוזומי בנות (או אחיות). של נוקלואידים שעדיין נמצאים בקשר הדוק זה עם זה. לאחר הפרדה כזו, הנוקלואידים מתפצלים ממרכז התא, ממקום מיקומם הקודם. יתרה מכך, אי התאמה זו מדויקת מאוד: רבע מאורך התא בשני כיוונים מנוגדים. כתוצאה מכך, שני נוקלואידים חדשים ממוקמים בתא. מה המנגנון לפער הזה? הוצע (Delamater, 1953) שחלוקת תאי חיידקים מקבילה למיטוזה אוקריוטית, אך במשך זמן רב לא היו ראיות התומכות בהנחה זו.
מידע חדש על מנגנוני חלוקת תאי חיידקים הושג על ידי חקר מוטנטים שבהם חלוקת התא הופרעה.
נמצאו כמה קבוצות של חלבונים מיוחדים מעורבים בתהליך ההפרדה הנוקלואידית. אחד מהם, חלבון Muk B, הוא הומודימר ענק (משקל מולקולרי כ-180 kDa, אורך 60 ננומטר), המורכב מחתך סליל מרכזי ומקטעים כדוריים סופיים, הדומים למבנה של חלבונים אוקריוטיים חוטיים (שרשרת מיוזין II, קינזין) . ב-N-terminus, Muk B נקשר ל-GTP ו-ATP, וב-C-terminus, למולקולת ה-DNA. תכונות אלו של Muk B נותנות סיבה לראות בו חלבון מוטורי המעורב בביקוע של נוקלואידים. מוטציות של חלבון זה מובילות להפרעות בהתבדלות הנוקלואידים: מספר רב של תאים נטולי גרעין מופיעים באוכלוסיית המוטנטים.
בנוסף לחלבון Muk B, צרורות של סיבים המכילים את חלבון Caf A, שיכולים להיקשר לשרשראות כבדות של מיוזין, כמו אקטין, משתתפות כנראה בהתבדלות הנוקלואידים (איור 331).
היווצרות של כיווץ, או מחיצה, גם ב במונחים כללייםדומה לציטוטומיה של תאי בעלי חיים. במקרה זה, חלבונים ממשפחת ה-Fts (רגישות תרמו פיברילרית) מעורבים ביצירת מחיצות. קבוצה זו כוללת מספר חלבונים, ביניהם החלבון FtsZ הוא הנחקר ביותר. זה דומה ברוב החיידקים, ארכיבקטריה, הוא נמצא ב-mycoplasmas וכלורופלסטים. זהו חלבון כדורי הדומה ברצף חומצות האמינו שלו לטובולין. בעת אינטראקציה עם GTP במבחנה, הוא מסוגל ליצור פרוטופילמנטים חוטיים ארוכים. ב-interphase, FtsZ ממוקם באופן דיפוזי בציטופלזמה, כמותו גדולה מאוד (5-20 אלף מונומרים לתא). במהלך חלוקת התא, כל החלבון הזה ממוקם באזור המחיצה, ויוצר טבעת מתכווצת, המזכירה מאוד את טבעת האקטומיוזין בחלוקה של תאי בעלי חיים (איור 332). מוטציות בחלבון זה מובילות להפסקת חלוקת התא: מופיעים תאים ארוכים המכילים נוקלואידים רבים. תצפיות אלו מראות תלות ישירה של חלוקת תאי חיידקים בנוכחות חלבוני Fts.
לגבי מנגנון היווצרות המחיצות, קיימות מספר השערות המניחות כיווץ של הטבעת באזור המחיצה, מה שמוביל לחלוקת התא המקורי לשניים. לפי אחד מהם, פרוטופילמנטים חייבים להחליק אחד ביחס לשני בעזרת חלבונים מוטוריים לא ידועים עדיין, לפי השני - הפחתה בקוטר המחיצה יכולה להתרחש עקב דה-פולימריזציה של FtsZ המעוגנת על קרום הפלזמה (איור 333).
במקביל להיווצרות המחיצה, נבנית שכבת ה-murein של דופן תא החיידק עקב עבודתו של הקומפלקס הפולי-אנזימטי PBP-3, המסנתז פפטידוגליקנים.
לפיכך, במהלך חלוקת תאי חיידקים מבוצעים תהליכים הדומים במידה רבה לחלוקה של אוקריוטים: התבדרות כרומוזומים (נוקלואידים) עקב אינטראקציה בין חלבונים מוטוריים ופיברילריים, היווצרות התכווצות עקב חלבונים פיברילרים. ליצור טבעת מתכווצת. בחיידקים, בניגוד לאאוקריוטים, חלבונים שונים לחלוטין לוקחים חלק בתהליכים אלה, אך עקרונות ארגון השלבים האישיים של חלוקת התא דומים מאוד.
שלח את העבודה הטובה שלך במאגר הידע הוא פשוט. השתמש בטופס למטה
סטודנטים, סטודנטים לתארים מתקדמים, מדענים צעירים המשתמשים בבסיס הידע בלימודיהם ובעבודתם יהיו אסירי תודה לכם מאוד.
חלוקת תאים מיטוטיים. ארגון כללי של מיטוזה
כפי שתיאוריית התאים מניחה, עלייה במספר התאים מתרחשת אך ורק בשל חלוקת התא המקורי, שהכפיל בעבר את החומר הגנטי שלו. זהו האירוע המרכזי בחיי התא כשלעצמו, כלומר השלמת השעתוק מסוגו. כל חיי ה"בין-פאזיים" של תאים מכוונים למימוש מלא של מחזור התא, המסתיים בחלוקת התא. חלוקת התא עצמה היא תהליך לא אקראי, שנקבע גנטית בהחלט, שבו שרשרת שלמה של אירועים מסודרת בשורה ברצף.
החלוקה של כל התאים האיקריוטים קשורה לעיבוי של כרומוזומים כפולים (משוכפלים), אשר לובשים צורה של מבנים חוטים צפופים. כרומוזומים חוטיים אלו מועברים לתאי בת על ידי מבנה מיוחד - ציר החלוקה. סוג זה של חלוקה של תאים אאוקריוטיים - מיטוזה (מיוונית מיטוס - חוטים), או קריוקינזיס, או חלוקה עקיפה - היא הדרך השלמה היחידה להגדיל את מספר התאים. חלוקת תאים ישירה או אמיטוזיס מתוארת בצורה מהימנה רק בחלוקת מקרו-גרעיניים פוליפלואידים של ריצות, המיקרו-גרעינים שלהם מתחלקים רק על ידי מיטוזה.
החלוקה של כל התאים האיקריוטים קשורה להיווצרות של מנגנון מיוחד לחלוקת תאים. במהלך שכפול התא מתרחשים שני אירועים: התבדרות כרומוזומים משוכפלים וחלוקת גוף התא, ציטוטומיה. החלק הראשון של האירוע באאוקריוטים מתבצע בעזרת מה שנקרא ציר החלוקה, המורכב ממיקרוטובולים, והחלק השני מתרחש עקב השתתפותם של מתחמי אקטומיוזין הגורמים להיווצרות התכווצות בתאים של בעלי חיים מקור או עקב השתתפותם של מיקרוטובולים וחוטי אקטין ביצירת פרגמופלסט, המחיצות התאיות הראשוניות בתאי הצמח.
שני סוגים של מבנים לוקחים חלק ביצירת ציר החלוקה בכל התאים האוקריוטיים: גופים קוטביים (קטבים) של הציר וקינטוכורים של כרומוזומים. גופים קוטביים, או צנטרוזומים, הם מרכזי הארגון (או הגרעין) של מיקרוטובולים. מיקרו-צינוריות צומחות מהן עם קצוות ה"+" שלהן, ויוצרות צרורות הנמתחות אל הכרומוזומים. בתאי בעלי חיים, הצנטרוזומים כוללים גם צנטרולים. אבל לאאוקריוטים רבים אין צנטריולים, ומרכזי ארגון מיקרו-צינוריות קיימים בצורה של אזורים אמורפיים חסרי מבנה, שמהם משתרעים מיקרו-טובוליות רבות. ככלל, שני צנטרוזומים או שני גופים קוטביים מעורבים בארגון מנגנון החלוקה, הממוקם בקצוות מנוגדים של גוף מורכב בצורת ציר המורכב ממיקרוטובולים. המבנה השני האופייני לחלוקת תאים מיטוטיים, המחבר את מיקרו-צינוריות הציר עם הכרומוזום, הם קינטוכורים. זה kinetochores, באינטראקציה עם microtubules, אשר אחראים על תנועת הכרומוזומים במהלך חלוקת התא.
סוגים שונים של מיטוזה איקריוטית
החלוקה של תאי בעלי חיים וצמחים שתוארה לעיל אינה הצורה היחידה של חלוקת תאים עקיפה. הסוג הפשוט ביותר של מיטוזה הוא pleuromitosis . במידה מסוימת הוא דומה לביקוע בינארי של תאים פרוקריוטים, שבו הנוקלואידים לאחר שכפול נשארים קשורים לממברנת הפלזמה, שמתחילה לצמוח כביכול בין נקודות הקישור של ה-DNA ובכך, כביכול, מפזרת את הכרומוזומים. לחלקים שונים של התא. לאחר מכן, במהלך היווצרות התכווצות התא, כל אחת ממולקולות ה-DNA תהיה בתא נפרד חדש.
כפי שכבר הוזכר, היווצרות ציר הבנוי ממיקרוטובולים אופיינית לחלוקת תאים אוקריוטיים. עם pleuromitosis סגורה (זה נקרא סגור בגלל ההתפצלות של כרומוזומים מתרחשת מבלי לשבור את הממברנה הגרעינית), לא centrioles, אלא מבנים אחרים הממוקמים בצד הפנימי של הממברנה הגרעינית, משתתפים כמרכזי ארגון מיקרו-צינוריות (MCMT). אלה הם מה שנקרא גופים קוטביים של מורפולוגיה בלתי מוגדרת, שמהם משתרעים מיקרוטובולים. ישנם שניים מהגופים הללו, הם מתפצלים זה מזה מבלי לאבד את הקשר שלהם עם המעטפת הגרעינית, וכתוצאה מכך נוצרים שני חצאי צירים הקשורים לכרומוזומים. כל תהליך היווצרות המנגנון המיטוטי וההתבדלות של הכרומוזומים מתרחשים במקרה זה מתחת לממברנה הגרעינית. סוג זה של מיטוזה נמצא בקרב פרוטוזואה, הוא נפוץ בפטריות (chytridia, zygomycetes, שמרים, oomycetes, ascomycetes, myxomycetes, וכו'). ישנן צורות של pleuromitosis סגורה למחצה, כאשר המעטפת הגרעינית נהרסת בקטבים של הציר שנוצר.
צורה נוספת של מיטוזה היא אורתומיטוזיס. במקרה זה, ה-COMTs ממוקמים בציטופלזמה; מההתחלה, לא נוצרים סמי-צירים, אלא ציר דו-קוטבי. ישנן שלוש צורות של אורתומיטוזיס: לִפְתוֹחַ(מיטוזה רגילה), חצי סגורו סָגוּר.באורתומיטוזיס סגורה למחצה, נוצר ציר ביסימטרי בעזרת TsOMT הממוקם בציטופלזמה, המעטפת הגרעינית נשמרת לאורך כל המיטוזה, למעט אזורי הקוטב. ניתן למצוא כאן המוני חומר גרגירי או אפילו צנטריולים בשם COMT. צורה זו של מיטוזה נמצאת באצות ירוקות, גרגרינים, אצות חומות, אצות אדומות ובכמה פטריות נמוכות יותר. עם אורתומיטוזיס סגור, הממברנה הגרעינית נשמרת לחלוטין, שמתחתיה נוצר ציר אמיתי. מיקרוטובולים נוצרים בקאריופלזמה, לעתים רחוקות יותר הם גדלים מ-COMT תוך גרעיני, שאינו קשור (בניגוד לפלאורומיטוזיס) עם הממברנה הגרעינית. סוג זה של מיטוזה מאפיין את חלוקתם של מיקרוגרעינים ריסים, אך נמצא גם בפרוטוזואים אחרים. באורתומיטוזיס פתוח, המעטפת הגרעינית מתפרקת לחלוטין. סוג זה של חלוקת תאים אופייני לאורגניזמים של בעלי חיים, כמה פרוטוזואה ותאים של צמחים גבוהים יותר. צורה זו של מיטוזה, בתורה, מיוצגת על ידי טיפוסים אסטרליים ואנסטרליים.
מתוך שיקול קצר זה ניתן לראות כי המאפיין העיקרי של מיטוזה באופן כללי הוא הופעת מבני ציר ביקוע, הנוצרים בקשר ל-CMTs, המגוונים במבנה שלהם.
מורפולוגיה של הדמות המיטוטית
כפי שכבר הוזכר, המנגנון המיטוטי נחקר באופן יסודי ביותר בתאים של צמחים ובעלי חיים גבוהים יותר. זה בא לידי ביטוי טוב במיוחד בשלב המטאפאזה של המיטוזה. בתאים חיים או קבועים במטאפאזה, במישור המשווני של התא, ממוקמים כרומוזומים, מהם מה שנקרא מתיחה בכיוונים מנוגדים. חוטי ציר המתכנסים בשני קטבים שונים של הדמות המיטוטית. אז הציר המיטוטי הוא אוסף של כרומוזומים, קטבים וסיבים. סיבי הציר הם מיקרוטובולים בודדים או צרורותיהם. מיקרו-צינוריות מתחילות מקטבי הציר וחלקן מגיעות לצנטרומרים, שם נמצאים ה-kinetochores של הכרומוזומים (kinetochore microtubules), חלקם הולכים יותר לכיוון הקוטב הנגדי, אך אינם מגיעים אליו - "מיקרוטובולים בין-קוטביים". בנוסף, קבוצה של מיקרוטובולים רדיאליים יוצאת מהקטבים, ויוצרת סביבם, כביכול, "זוהר קורן" - אלה הם מיקרוטובולים אסטרליים.
על פי המורפולוגיה הכללית, דמויות מיטוטיות מחולקות לשני סוגים: אסטרלי ואנסטרלי.
סוג הציר האסטרלי (או מתכנס) מאופיין בכך שהקטבים שלו מיוצגים על ידי אזור קטן שאליו מתכנסות (מתכנסות) המיקרוטובולים. בדרך כלל, צנטרוזומים המכילים צנטרולים ממוקמים בקטבים של הצירים האסטרליים. אמנם ידועים מקרים של מיטוזות אסטרליות צנטרולריות (במהלך מיוזה של חלק מחסרי חוליות). בנוסף, מיקרוטובולים רדיאליים מתפצלים מהקטבים, שאינם חלק מהציר, אלא יוצרים אזורי כוכבים - ציסטרים. באופן כללי, סוג זה של ציר מיטוטי הוא יותר כמו משקולת.
לסוג האנסטריאלי של הדמות המיטוטית אין ציסטרים בקטבים. האזורים הקוטביים של הציר כאן רחבים, הם נקראים כובעים קוטביים, הם לא כוללים centrioles. סיבי הציר במקרה זה אינם יוצאים מנקודה אחת, אלא מתפצלים בחזית רחבה (מתפצלים) מכל אזור כובעי הקוטב. סוג זה של ציר מאפיין תאים מתחלקים של צמחים גבוהים יותר, אם כי הוא נמצא לעתים בבעלי חיים גבוהים יותר. כך, למשל, בעוברות מוקדמת של יונקים במהלך חלוקת הבשלת הביציות ובמהלך חלוקות I ו-II של הזיגוטה, נצפו מיטוזות צנטרולריות (מתפצלות). אבל החל מחלוקת התא השלישית ובכל אלה שלאחר מכן, תאים מתחלקים בהשתתפות צירים אסטרליים, שבקטבים שלהם נמצאים תמיד צנטרולים.
באופן כללי, עבור כל צורות המיטוזה, הכרומוזומים עם הקינטוכורים שלהם, הגופים הקוטביים (צנטרוזומים) וסיבי הציר נשארים מבנים נפוצים.
דינמיקה של מיטוזיס
בתאים שנכנסו למחזור החלוקה, שלב המיטוזה עצמו, החלוקה העקיפה, לוקח זמן קצר יחסית, רק כ-0.1 מזמן מחזור התא. אז, בתאים מתחלקים של מריסטם השורש, האינטרפאזה יכולה להיות 16-30 שעות, והמיטוזה יכולה להימשך רק 1-3 שעות. המחזור של תאי אפיתל מעיים של עכבר נמשך כ-20-22 שעות, בעוד שהמיטוזה נמשכת רק שעה. ריסוק ביצים, כל תקופת התא, כולל מיטוזה, עשויה להיות פחות משעה.
תהליך חלוקת התאים המיטוטי מחולק בדרך כלל למספר שלבים עיקריים: פרופאזה, פרומטפאזה, מטאפאזה, אנפאזה, טלופזה. קשה מאוד לקבוע את הגבולות בין השלבים הללו במדויק, כי המיטוזה עצמה היא תהליך מתמשך ושינוי השלבים מתרחש בהדרגה רבה: אחד מהם עובר באופן בלתי מורגש לאחר. השלב היחיד שיש לו התחלה אמיתית הוא אנפאזה - תחילת תנועת הכרומוזומים לכיוון הקטבים. משך שלבים בודדים של מיטוזה שונה, הקצר ביותר בזמן הוא אנפאזה (טבלה).
משך שלבי המיטוזה
הזמן של שלבים בודדים של מיטוזה נקבע בצורה הטובה ביותר על ידי התבוננות ישירה של חלוקת תאים חיים בחדרים מיוחדים. לדעת את זמן המיטוזה, אפשר לחשב את משך שלבים בודדים לפי אחוז התרחשותם בין תאים מתחלקים.
שלבים של מיטוזה
פרופאזה.כבר בסוף תקופת G 2 מתחילים להתרחש סידורים משמעותיים מחדש בתא. אי אפשר לקבוע בדיוק מתי מתרחשת הפרופזה. הקריטריון הטוב ביותר לתחילת שלב זה של מיטוזה יכול להיות ההופעה בגרעינים של מבנים חוטים - כרומוזומים מיטוטיים. לאירוע זה מקדימה עלייה בפעילות של פוספורילאזים המשנים היסטונים, וקודם כל, היסטון H1. בפרופאזה, כרומטידות אחיות מקושרות זו לצד זו בעזרת חלבוני קוהזין, היוצרים את הקשרים הללו כבר בתקופת S, במהלך שכפול הכרומוזומים. לפי הפרופזה המאוחרת, הקשר בין כרומטידות אחיות נשמר רק באזור הקינטוכורים. בכרומוזומי פרופאזה כבר ניתן לראות קינטוכורים בוגרים, שאין להם קשרים עם מיקרוטובולים.
עיבוי הכרומוזומים בגרעין הפרופזה חופף לירידה חדה בפעילות התעתיק של הכרומטין, שנעלם לחלוטין באמצע הפרופזה. עקב ירידה בסינתזה של RNA ועיבוי כרומטין, מתרחשת גם נטרול של גנים גרעיניים. במקביל, מרכזי פיברילרים בודדים מתמזגים כך שהם הופכים למקטעים יוצרים גרעין של כרומוזומים, למארגנים גרעיניים. רוב החלבונים הגרעיניים מתנתקים ונמצאים בצורה חופשית בציטופלזמה של התא או נקשרים לפני השטח של הכרומוזומים.
במקביל מתרחשת זרחון של מספר חלבונים של הלמינה, הממברנה הגרעינית, המתפרקת. במקרה זה אבד הקשר של המעטפת הגרעינית עם הכרומוזומים. ואז המעטפת הגרעינית מפוצלת ל-vacuoles קטנים, ומתחמי הנקבוביות נעלמים.
במקביל לתהליכים אלו מתרחשת הפעלה של מרכזי תאים. בתחילת הפרופזה, המיקרוטובולים בציטופלזמה מתפרקים ומתחילה צמיחה מהירה של מיקרוטובולים אסטרליים רבים סביב כל אחד מהדיפלוזומים הכפולים. קצב הגדילה של מיקרוטובוליות בפרופאזה גבוה כמעט פי שניים מהצמיחה של מיקרוטובוליות בין-פאזיות, אך רגישותן גבוהה פי 5-10 מזו של ציטופלזמה. אז אם זמן מחצית החיים של המיקרוטובולים בציטופלזמה הוא בערך 5 דקות, אז במהלך המחצית הראשונה של המיטוזה הוא רק 15 שניות. כאן, חוסר היציבות הדינמית של המיקרוטובולים בולטת עוד יותר. כל המיקרוטובולים המשתרעים מהצנטרוזומים גדלים קדימה עם הקצוות (+).
צנטרוזומים פעילים - עמודי הציר העתידיים - מתחילים להתפצל זה מזה למרחק מסוים. המנגנון של התבדלות פרופאזה כזו של הקטבים הוא כדלקמן: מיקרוטובולים אנטי-מקבילים הנעים זה לעבר זה מקיימים אינטראקציה זה עם זה, מה שמוביל לייצוב ולדחייה גדולים יותר של הקטבים. זה מתרחש עקב האינטראקציה עם מיקרו-צינוריות של חלבונים דמויי דיניין, אשר בחלק המרכזי של הציר מסדרים מיקרו-צינוריות בין-קוטביות במקביל זו לזו. במקביל נמשכים הפילמור והגדילה שלהם, המלווים במקביל לדחיפתם לכיוון הקטבים עקב עבודתם של חלבונים דמויי קינזין. בשלב זה, במהלך היווצרות הציר, המיקרוטובולים עדיין אינם קשורים לקינטוכורים של הכרומוזומים.
בפרופאזה, במקביל לפירוק המיקרוטובולים הציטופלזמיים, הרטיקולום האנדופלזמי אינו מאורגן (הוא מתפרק לוואקווולים קטנים השוכנים לאורך פריפריית התא) ומנגנון גולגי, שמאבד את הלוקליזציה הפרי-גרעינית שלו, מתפרק לדיקטיוזומים נפרדים המפוזרים באקראי בציטופלזמה. .
פרומטפאזה.לאחר הרס המעטפת הגרעינית, הכרומוזומים המיטוטים שוכנים באזור הגרעין הקודם ללא סדר מסוים. בפרומטפאזה מתחילה תנועתם ותנועתם, מה שיוביל בסופו של דבר להיווצרות "צלחת" של כרומוזומים משווני, לסידור מסודר של כרומוזומים בחלק המרכזי של הציר שכבר נמצא במטאפאזה. בפרומטפאזה, ישנה תנועה מתמדת של כרומוזומים או מטאפאזה, שבה הם מתקרבים אל הקטבים, או משאירים אותם לכיוון מרכז הציר עד שהם תופסים את המיקום האמצעי האופייני למטאפאזה (קונגרסיה של כרומוזומים).
בתחילת הפרומטאפאזה, הכרומוזומים השוכנים קרוב יותר לאחד הקטבים של הציר שנוצר מתחילים להתקרב אליו במהירות. זה לא קורה בבת אחת, אבל לוקח קצת זמן. נמצא שסחיפה אסינכרונית ראשונית כזו של כרומוזומים לקטבים שונים מתרחשת בעזרת מיקרוטובולים. באמצעות שיפור ניגודיות פאזה וידאו-אלקטרונית במיקרוסקופ אור, ניתן היה לראות על תאים חיים שמיקרוטובולים בודדים המשתרעים מהקטבים מגיעים בטעות לאחד מהקינטוכורים של הכרומוזום ונקשרים אליו, "נלכדים" על ידי הקינטוכור. לאחר מכן, ישנו כרומוזום מהיר, במהירות של כ-25 מיקרומטר לדקה, גלישת כרומוזומים לאורך המיקרוטובול לכיוון (-)-קצהו. זה גורם לכרומוזום להתקרב אל הקוטב שממנו הגיעה המיקרוטובולה הזו. חשוב לציין כי kinetochores יכולים ליצור קשר עם פני השטח לרוחב של microtubules כאלה. במהלך תנועה זו של הכרומוזומים, המיקרוטובולים אינם מפורקים. סביר להניח שחלבון מוטורי הדומה לדינאין הציטופלזמי המצוי בכתר הקינטוכורים אחראי לתנועה כה מהירה של כרומוזומים.
כתוצאה מתנועת פרומטפאזה ראשונית זו, הכרומוזומים מתקרבים באופן אקראי אל קטבי הציר, שם נמשכת היווצרותם של מיקרוטובולים חדשים. ברור שככל שהקינטוכור הכרומוזומלי קרוב יותר לצנטרוזום, כך גדלה האקראיות של האינטראקציה שלו עם מיקרוטובולים אחרים. במקרה זה, קצוות חדשים וגדלים (+) של מיקרוטובולים "נלכדים" על ידי אזור הכתר הקינטוכור; כעת מחוברת צרור של מיקרוטובוליות לקינטוכור, שצמיחתן ממשיכה בקצה (+) שלהן. עם צמיחתו של צרור כזה, הקינטוכור, ואיתו הכרומוזום, חייבים לנוע לכיוון מרכז הציר, להתרחק מהקוטב. אבל בשלב זה, מיקרוטובולים צומחים מהקוטב הנגדי לקינטוכור השני של הכרומטיד האחות השני, שהצרור שלו מתחיל למשוך את הכרומוזום לקוטב הנגדי. נוכחותו של כוח משיכה כזה מוכחת על ידי העובדה שאם חותכים צרור של מיקרו-צינוריות באחד מהקינטוכורים באמצעות מיקרו-קרן לייזר, הכרומוזום מתחיל לנוע לכיוון הקוטב הנגדי. בתנאים רגילים, הכרומוזום, המבצע תנועות קטנות לכיוון הקוטב האחד או השני, כתוצאה מכך תופס בהדרגה מיקום אמצעי בציר. בתהליך של סחף פרומטפאזה של כרומוזומים, המיקרוטובוליות מתארכות ונבנות בקצוות (+) כאשר הקינטוכור מתרחק מהקוטב, ומיקרוטובולים מפורקים ומתקצרים גם בקצה (+), כאשר הקינטוכור האחות. נע לכיוון הקוטב.
תנועות מתחלפות אלו של הכרומוזומים קדימה ואחורה מובילות לעובדה שהם בסופו של דבר מגיעים לקו המשווה של הציר ומסתדרים בשורה בלוח המטאפאזה.
מטאפאזה. במטאפאזה, כמו גם בשלבים אחרים של מיטוזה, למרות ייצוב מסוים של צרורות המיקרוטובוליות, חידושם המתמיד נמשך עקב הרכבה ופירוק של טובולינים. במהלך המטאפאזה, הכרומוזומים מסודרים כך שהקינטוכורים שלהם פונים לקטבים מנוגדים. יחד עם זאת, יש מחיצה קבועה ומיקרוטובולים בין-קוטביים, שמספרם במטאפאזה מגיע למקסימום. אם מסתכלים על תא המטאפאזה מהצד של הקוטב, אז אפשר לראות שהכרומוזומים מסודרים כך שהקטעים הצנטרומריים שלהם פונים למרכז הציר, והכתפיים פונות לפריפריה. סידור זה של כרומוזומים נקרא "כוכב האם" והוא אופייני לתאי בעלי חיים. בצמחים, לעתים קרובות במטאפאזה, הכרומוזומים שוכנים במישור המשווני של הציר ללא סדר קפדני.
בסוף המטאפאזה, תהליך ההפרדה של כרומטידות אחיות זו מזו הושלם. הכתפיים שלהם מקבילות זו לזו, הפער המפריד ביניהם נראה בבירור ביניהן. המקום האחרון שבו נשמר המגע בין הכרומטידות הוא הצנטרומר; עד סוף המטאפאזה, הכרומטידות בכל הכרומוזומים נשארות מחוברות באזורים הצנטרומריים.
אנפאזהמתחיל בפתאומיות, שניתן לראות היטב במחקר החיוני. אנפאזה מתחילה בהפרדה של כל הכרומוזומים בבת אחת באזורים הצנטרומריים. בזמן זה, ישנה השפלה בו-זמנית של קוהזינים צנטרומרים, שעד לאותו זמן קשרו כרומטידות אחיות. הפרדה בו-זמנית זו של הכרומטידים מאפשרת להם להתחיל את ההפרדה הסינכרונית שלהם. כרומוזומים מאבדים לפתע את הרצועות הצנטרומריות שלהם ומתחילים להתרחק זה מזה באופן סינכרוני לעבר קטבים מנוגדים של הציר. מהירות תנועת הכרומוזומים אחידה, היא יכולה להגיע ל-0.5-2 מיקרומטר/דקה.
אנפאזה היא השלב הקצר ביותר של מיטוזה (כמה אחוזים מהזמן הכולל), אך במהלך תקופה זו מתרחשים מספר אירועים. העיקריים שבהם הם הפרדה של שתי קבוצות זהות של כרומוזומים והובלתם לקצוות מנוגדים של התא.
כאשר הכרומוזומים נעים, הם משנים את הכיוון שלהם ולעתים קרובות מקבלים צורת V. החלק העליון שלהם מופנה לכיוון מוטות החלוקה, והכתפיים נזרקות, כביכול, חזרה למרכז הציר. אם שבר בזרוע הכרומוזומים התרחש לפני אנפאזה, אז במהלך האנפאזה הוא לא ישתתף בתנועת הכרומוזומים ויישאר באזור המרכזי. תצפיות אלו הראו שהאזור הצנטרומרי, יחד עם הקינטוכור, הוא זה שאחראי לתנועת הכרומוזומים. נראה שהכרומוזום נמשך אל הקוטב שמעבר לצנטרומר. בחלק מהצמחים הגבוהים יותר (אוסיקה) אין התכווצות צנטרומרית בולטת, וסיבי הציר נמצאים במגע עם נקודות רבות על פני הכרומוזומים (כרומוזומים פולצנטריים והולוצנטריים). במקרה זה, הכרומוזומים ממוקמים על פני סיבי הציר.
למעשה, הפרדת כרומוזומים מורכבת משני תהליכים: 1 - הפרדת כרומוזומים עקב צרורות קינטוכורים של מיקרוטובולים, 2 - הפרדת כרומוזומים יחד עם קטבים עקב התארכות של מיקרוטובולים בין-קוטביים. הראשון מבין התהליכים הללו נקרא "אנפאזה A", השני - "אנפאזה B".
במהלך אנפאזה A, כאשר קבוצות של כרומוזומים מתחילות לנוע לכיוון הקטבים, מתרחש קיצור של צרורות הקינטוכור של המיקרוטובולים. ניתן היה לצפות שבמקרה זה דה-פולימריזציה של מיקרוטובולים תתרחש בקצוות (-) שלהן, הקצוות הקרובים ביותר לקוטב. עם זאת, הוכח שמיקרוטובולים אכן מפורקים, אך בעיקר (80%) מהקצוות (+) הסמוכים לקינטוכורים. בניסוי, טובולין הקשור ל-fluorochrome הוכנס לתאי תרבית רקמה חיים בשיטת המיקרו-הזרקה. זה איפשר לראות באופן חיוני את המיקרוטובולים בציר הביקוע. בתחילת האנפאזה, צרור הציר של אחד הכרומוזומים הוקרן במיקרו-קרן קלה בערך באמצע בין הקוטב לכרומוזום. עם חשיפה זו, הקרינה נעלמת באזור המוקרן. תצפיות הראו שהאזור המוקרן אינו מתקרב אל הקוטב, אך הכרומוזום מגיע אליו כאשר צרור הקינטוכור מתקצר. כתוצאה מכך, פירוק המיקרוטובולים של צרור הקינטוכור מתרחש בעיקר מהקצה (+) בנקודת החיבור שלו עם הקינטוכור, והכרומוזום נע לכיוון הקצה (-) של המיקרוטובולים, שנמצא ב-. אזור הצנטרוזום. התברר שתנועה כזו של כרומוזומים תלויה בנוכחות ATP ובנוכחות של ריכוז מספיק של יוני Ca+. העובדה שחלבון הדיניין נמצא בהרכב הכתר הקינטוכור, שבו משובצים קצוות ה-(+) של המיקרוטובולים, אפשרה לנו לשקול שחלבון זה הוא שמושך את הכרומוזום אל הקוטב. במקביל, מתרחשת דה-פולימריזציה של microtubules kinetochore בקצה (+).
לאחר הפסקת הכרומוזומים בקטבים, מתרחשת התבדלות נוספת שלהם עקב הוצאת הקטבים זה מזה (אנפאזה B). הוכח שבמקרה זה, הקצוות (+) של המיקרוטובולים הבין-קוטביים גדלים, אשר יכולים להגדיל משמעותית את אורכם. האינטראקציה בין המיקרוטובולים האנטי-מקבילים הללו, המובילה להחלקתן ביחס זו לזו, נקבעת על ידי חלבונים דמויי קינזין מוטוריים אחרים. בנוסף, הקטבים נמשכים בנוסף לפריפריה של התא עקב אינטראקציה של חלבונים דמויי דיניין על קרום הפלזמה עם מיקרוטובולים אסטרליים.
רצף האנפאזות A ו-B ותרומתם לתהליך הפרדת הכרומוזומים יכולים להיות שונים באובייקטים שונים. אז, ביונקים, שלבים A ו-B מתרחשים כמעט בו זמנית. בפרוטוזואה B, אנפאזה יכולה לגרום לעלייה של פי 15 באורך הציר. שלב ב' נעדר בתאי הצמח.
טלופאזמתחיל בעצירת כרומוזומים (טלופאזה מוקדמת, אנפאזה מאוחרת) ומסתיים עם תחילת שחזור של גרעין בין-פאזי חדש (תקופת G 1 מוקדמת) וחלוקת התא המקורי לשני תאי בת (ציטוקינזיס).
בטלופאזה המוקדמת, הכרומוזומים, מבלי לשנות את כיוונם (אזורים צנטרומריים - לכיוון הקוטב, אזורים טלומריים - לכיוון מרכז הציר), מתחילים להתעבות ולהגדיל את נפחם. באתרי המגע שלהם עם שלפוחיות ממברנות של הציטופלזמה, מתחיל להיווצר קרום גרעיני חדש, שנוצר תחילה על המשטחים הצדדיים של הכרומוזומים ובהמשך באזורים הצנטרומריים והטלומריים. לאחר סגירת הממברנה הגרעינית, מתחילה היווצרותם של נוקלאולים חדשים. התא נכנס לתקופת G 1 של שלב ביניים חדש.
בטלופאזה מתחיל ונגמר תהליך ההרס של המנגנון המיטוטי - פירוק המיקרוטובולים. הוא עובר מהקטבים לקו המשווה של התא הקודם: זה בחלק האמצעי של הציר שהמיקרוטובולים נמשכים הכי הרבה זמן (שיורי גוף).
אחד האירועים העיקריים של טלופאז הוא חלוקת גוף התא, ציטוטומיהאוֹ ציטוקינזיס.כבר נאמר לעיל כי בצמחים, חלוקת תאים מתרחשת על ידי היווצרות תוך תאית של מחיצת תאים, ובתאים של בעלי חיים, על ידי התכווצות, החדרה של קרום הפלזמה לתוך התא.
מיטוזה לא תמיד מסתיימת בחלוקת גוף התא. לפיכך, באנדוספרם של צמחים רבים, תהליכים מרובים של ביקוע גרעיני מיטוטי יכולים להתרחש במשך זמן מה ללא חלוקה ציטופלזמית: נוצר סימפלסט רב-גרעיני ענק. כמו כן, ללא ציטוטומיה, גרעינים רבים של פלסמודיה של מיקומיציטים מתחלקים באופן סינכרוני. בשלבים המוקדמים של התפתחות העוברים של חרקים מסוימים, ביקוע חוזר של גרעינים מתרחש גם ללא חלוקה של הציטופלזמה.
ברוב המקרים, היווצרות ההיצרות במהלך חלוקת תאי בעלי חיים מתרחשת אך ורק במישור המשווני של הציר. כאן, בסוף האנפאזה, בתחילת הטלופזה, נוצרת הצטברות קליפת המוח של מיקרופילמנטים היוצרים טבעת מתכווצת. המיקרופילמנטים של הטבעת כוללים סיבי אקטין ומולקולות קצרות בצורת מוט של מיוזין II מפולמר. החלקה הדדית של רכיבים אלו מובילה לירידה בקוטר הטבעת ולהופעת חריץ של קרום הפלזמה, מה שמוביל לבסוף להתכווצות התא המקורי לשניים.
לאחר ציטוטומיה, שני תאים חדשים (בת) נכנסים לשלב G 1 של תקופת התא. בשלב זה, הסינתזות הציטופלזמיות מתחדשות, המערכת הוואקואולרית משוחזרת, הדיקטיוזומים של מנגנון גולגי מתרכזים שוב באזור הפרי-גרעיני בשיתוף עם הצנטרוזום. מהצנטרוזום מתחילה צמיחתם של מיקרוטובולים ציטופלזמיים ושיקום הציטושלד הבין-פאזי.
מיטוזה של תאי צמח
לחלוקת תאים מיטוטיים של צמחים גבוהים יש מספר מאפיינים אופייניים המתייחסים לתחילתו ולסופו של תהליך זה.
בתאי interphase של מריסטמים צמחיים שונים, microtubules ממוקמים בשכבת תת-ממברנה קליפת המוח של הציטופלזמה, ויוצרים צרורות טבעתיים של microtubules. מיקרוטובולים היקפיים נמצאים במגע עם אנזימים היוצרים סיבים תאית, עם סינתטאז של תאית, שהם חלבונים אינטגרליים של קרום הפלזמה. הם מסנתזים תאית על פני ממברנת הפלזמה. מאמינים שבמהלך הצמיחה של סיב התאית, אנזימים אלה נעים לאורך המיקרוטובולים התת-ממברניים.
סידור מחדש מיטוטי של אלמנטים ציטו-שלד מתרחש בתחילת הפרופזה. במקביל, מיקרוטובוליות נעלמות בשכבות ההיקפיות של הציטופלזמה, אך בשכבה הקרובה לממברנה של הציטופלזמה באזור קו המשווה של התא מופיעה צרור טבעתי של מיקרוטובולים - טבעת קדם-פרופאזית, הכוללת יותר מ-100 מיקרוטובולים. . מבחינה אימונוכימית, גם בטבעת זו נמצא אקטין. חשוב לציין שהטבעת הקדם-פרופאזית של המיקרוטובולים ממוקמת במקום בו תיווצר מחיצת תאים בטלופאז המפרידה בין שני תאים חדשים. מאוחר יותר בפרופזה, הטבעת הזו מתחילה להיעלם, ומיקרוטובוליות חדשות מופיעות לאורך הפריפריה של גרעין הפרופזה. מספרם גדול יותר באזורי הקוטב של הגרעינים; הם, כביכול, עוטפים את כל הפריפריה הגרעינית. במהלך המעבר ל-prometaphase, מתעורר ציר דו-קוטבי, שהמיקרוטובולים שלו מתקרבים למה שנקרא. כובעים קוטביים, שבהם נצפים רק ואקוולים קטנים וסיבים דקים בעלי מורפולוגיה לא ודאית; לא נמצאו סימנים של צנטריולים באזורי הקוטב הללו. כך נוצר הציר האנאסטרלי.
בפרומטפאזה, במהלך חלוקת תאי הצמח, נצפית גם סחיפה מורכבת של כרומוזומים, תנודה ותנועה שלהם מאותו סוג המתרחשות בפרומטפאזה של תאי בעלי חיים. אירועים באנפאזה דומים גם לאלו במיטוזה אסטרלית. לאחר התפצלות הכרומוזומים, נוצרים גרעינים חדשים, גם עקב פירוק הכרומוזומים והיווצרות מעטפת גרעינית חדשה.
תהליך הציטוטומיה של תאי צמחים שונה באופן חד מחלוקת התכווצות של תאי בעלי חיים. במקרה זה, פירוק של microtubules ציר באזורי הקוטב מתרחש גם בסוף הטלופזה. אבל המיקרו-צינוריות של החלק העיקרי של הציר בין שני הגרעינים החדשים נשארות, יתר על כן, היווצרותם של מיקרו-צינוריות חדשות מתרחשת כאן. כך נוצרות צרורות של מיקרו-צינוריות, שאליהן משויכים תאים קטנים רבים. ואקוולים אלו מקורם בוואקואולים של מנגנון גולגי ומכילים חומרים פקטין. בעזרת מיקרו-צינוריות עוברים וואקוולים רבים לאזור המשווה של התא, שם הם מתמזגים זה עם זה ויוצרים ואקוול שטוח באמצע התא - פרגמופלסט שצומח לכיוון פריפרית התא, כולל עוד ועוד. ואקוולים חדשים.
כך נוצר דופן התא הראשוני. בסופו של דבר, קרומי הפרגמופלסט מתמזגים עם קרום הפלזמה: שני תאים חדשים נפרדים, מופרדים על ידי דופן תא חדש שנוצר. ככל שהפרגמופלסט מתרחב, צרורות המיקרוטובוליות נעות יותר ויותר לעבר פריפריית התא. סביר להניח שתהליך מתיחה של הפרגמופלסט והעברת צרורות המיקרוטובולים לפריפריה מקל על ידי צרורות של חוטי אקטין הנמשכים מהשכבה הקורטיקלית של הציטופלזמה במקום שבו הייתה טבעת הקדם-פרופאזה.
לאחר חלוקת התא, המיקרוטובוליות המעורבות בהובלת ואקוולים קטנים נעלמים. דור חדש של microtubules interphase נוצר על הפריפריה של הגרעין, ולאחר מכן ממוקם בשכבת הממברנה הקורטיקלית של הציטופלזמה.
זהו תיאור כללי של חלוקת תאי צמחים, אך תהליך זה אינו מובן בצורה גרועה ביותר. באזורי הקוטב של הצירים לא נמצאו חלבונים שהם חלק מה-COMT של תאי בעלי חיים. נמצא שבתאי צמחים ניתן למלא תפקיד זה על ידי הממברנה הגרעינית, שממנה מופנים קצוות ה-(+) של המיקרו-צינוריות אל פריפריית התא, ואת הקצוות (-) אל הממברנה הגרעינית. כאשר הציר נוצר, צרורות הקינטוכורים מכוונים עם קצה (-) שלהם אל המוט, וקצה (+) אל הקינטוכורים. כיצד מתרחש כיוון מחדש זה של מיקרוטובולים עדיין לא ברור.
במהלך המעבר לפרופזה, מופיעה מסביב לגרעין רשת צפופה של מיקרו-צינוריות, הדומה לסל, אשר לאחר מכן מתחילה להידמות לציר בצורתו. במקרה זה, מיקרוטובולים יוצרים סדרה של צרורות מתכנסים המכוונים לקטבים. מאוחר יותר בפרומטאפאזה מתרחש הקשר של מיקרוטובולים עם קינטוכורים. במטאפאזה, סיבים של קינטוכור יכולים להוות מרכז התכנסות משותף - מיני קוטבי ציר, או מרכזי התכנסות של מיקרוטובולים. סביר להניח, היווצרות של minipoles כאלה מתרחשת עקב האיחוד של הקצוות (-) של microtubules הקשורים kinetochores. ניתן להניח שבתאי הצמחים הגבוהים, תהליך הארגון מחדש של השלד הציטוטי, כולל היווצרות הציר המיטוטי, קשור לארגון העצמי של המיקרוטובולים, אשר, כמו בתאי בעלי חיים, מתרחש בהשתתפותם של חלבונים מוטוריים.
מסמכים דומים
חקר תהליך המיטוזה כחלוקת תאים עקיפה ושיטת רבייה נפוצה של תאים אוקריוטיים, משמעותו הביולוגית. מיוזיס היא חלוקת תא הפחתה. אינטרפאזה, פרופאזה, מטאפאזה, אנפאזה וטלופאזה של מיוזה ומיטוזה.
מצגת, נוספה 21/02/2013
חלוקת תאים מיטוטיים, תכונות המבנה שלו. מיטוזיס כשיטה אוניברסלית לחלוקת תאים בצמחים ובעלי חיים. הקביעות של המספר והאינדיבידואליות של הכרומוזומים. תוחלת חיים, הזדקנות ומוות תאים. צורות רבייה של אורגניזמים.
תקציר, נוסף 07.10.2009
משמעות הצמיחה וההתפתחות של תאים. חיים ומחזורים מיטוטיים של תאים. תוחלת חיים של סוגים שונים של תאים באורגניזם רב תאי. התחשבות במיטוזה כשיטת רבייה אוניברסלית, המשמרת את הקביעות של מספר הכרומוזומים בתאים.
מצגת, נוספה 12/05/2014
השלבים העיקריים של מחזור התא: אינטרפאזה ומיטוזה. הגדרת המושג "מיטוזה" כחלוקת תאים עקיפה, השיטה הנפוצה ביותר להתרבות של תאים אוקריוטיים. מאפיינים ומאפיינים של תהליכי חלוקה: אמיטוזיס ומיוזה.
מצגת, נוספה 25/10/2011
הגרעין של תא איקריוטי. תאים שיש להם יותר משתי קבוצות של כרומוזומים. תהליך החלוקה באוקריוטים. זוגות מאוחדים כרומוזומים הומולוגיים. אונטוגניה של תאי צמחים. תהליך הפרדת התאים כתוצאה מהרס של הלמינה החציונית.
תקציר, נוסף 28/01/2011
מיטוזיס היא חלוקת תאים עקיפה שגורמת להיווצרות תאים סומטיים. שלבי מחזור התא. הכנה לחלוקת אורגניזמים אוקריוטיים. השלבים העיקריים של קריוקינזיס. הפרדת ציטופלזמה עם אברונים בין תאי בת.
מצגת, נוספה 11/06/2013
מרכיבים של תא צמחי. קרום פלזמה ותפקידיה. רכיבי דופן התא. סוגי מיטוזה באאוקריוטים. רקמות חינוכיות בגוף הצמחים ומיקומן. תכונות מכאניותתאי צמחים. רקמות הפרשה חיצוניות.
מדריך, נוסף ב-12/12/2009
מאפייני מחזור החיים של תא, מאפיינים של תקופות קיומו מחלוקה לחלוקה הבאה או למוות. שלבי המיטוזה, משך הזמן שלהם, טבעם ותפקיד האמיטוזה. המשמעות הביולוגית של המיוזה, שלביה וזנייה העיקריים.
הרצאה, נוספה 27/07/2013
תקופות ושלבים של מחזור התא. מעבר רציף של תקופות המחזור על ידי התא מבלי לדלג או לחזור לשלבים הקודמים. חלוקת התא המקורי לשני תאי בת. ציקלין וקינאזות תלויות ציקלין; חלוקת תאים אוקריוטיים; מיטוזה.
עבודת בקרה, נוסף 21/11/2009
הסוגים העיקריים של תאים חיים ותכונות המבנה שלהם. תכנית כלליתמבנים של תאים איקריוטים ופרוקריוטיים. תכונות של המבנה של תאי צמחים ופטריות. טבלת השוואהמבני תאים של צמחים, בעלי חיים, פטריות וחיידקים.
קיימות שתי שיטות חלוקה: 1) החלוקה השכיחה ביותר, השלמה - מיטוזה (חלוקה עקיפה) ו-2) אמיטוזיס (חלוקה ישירה). במהלך החלוקה המיטוטית, הציטופלזמה נבנית מחדש, המעטפת הגרעינית נהרסת ומזהים כרומוזומים. בחייו של תא ישנה תקופה של מיטוזה עצמה ומרווח בין חלוקות, הנקרא interphase. עם זאת, תקופת האינטרפאזה (תאים שאינם מתחלקים) במהותה יכולה להיות שונה. במקרים מסוימים, במהלך האינטרפאזה, התא מתפקד ובו זמנית מתכונן לחלוקה הבאה. במקרים אחרים, תאים נכנסים ל-interphase, מתפקדים, אך אינם מתכוננים עוד לחלוקה. כחלק מאורגניזם רב-תאי מורכב, ישנן קבוצות רבות של תאים שאיבדו את היכולת להתחלק. אלה כוללים, למשל, תאי עצב. הכנת תאים למיטוזה מתרחשת ב-interphase. על מנת לדמיין את המאפיינים העיקריים של תהליך זה, זכור את המבנה של גרעין התא.
תאי בצל בשלבים שונים של מחזור התא
היחידה המבנית הבסיסית של הגרעין היא הכרומוזומים, המורכבים מ-DNA וחלבון. בגרעינים של תאים חיים שאינם מתחלקים, ככלל, לא ניתן להבחין בכרומוזומים בודדים, אך רוב הכרומטין, שנמצא על תכשירים מוכתמים בצורת חוטים דקים או גרגרים בגדלים שונים, מתאים לכרומוזומים. בתאים מסוימים, כרומוזומים בודדים נראים היטב גם בגרעין הבין-פאזי, למשל, בתאים המתחלקים במהירות של ביצית מופרית מתפתחת ובגרעינים של פרוטוזואה מסויימת. בְּ תקופות שונותבמהלך חיי התא, הכרומוזומים עוברים שינויים מחזוריים שניתן לעקוב אחריהם מחלוקה אחת לאחרת. כרומוזומים בזמן מיטוזה הם גופים צפופים מוארכים, שלאורכם ניתן להבחין בשני גדילים - כרומטידות המכילות DNA, שהן תוצאה של הכפלת הכרומוזומים. לכל כרומוזום יש כיווץ ראשוני, או צנטרומר. חלק מצומצם זה של הכרומוזום יכול להיות ממוקם באמצע או קרוב יותר לאחד הקצוות, אבל עבור כל כרומוזום מסוים מקומו קבוע בהחלט. במהלך מיטוזה, הכרומוזומים והכרומטידים הם חוטים סליליים מפותלים בחוזקה (מצב ספירלי או מעובה). בגרעין הבין-פאזי, הכרומוזומים מוארכים מאוד, כלומר, מפושטים, עקב כך קשה להבחין ביניהם. כתוצאה מכך, מחזור השינויים בכרומוזומים מורכב מספירליזציה, כאשר הם מתקצרים, מתעבים ונעשים ניתנים להבחנה בבירור, ו-despiralization, כאשר הם מוארכים מאוד, שזורים זה בזה, ואז זה הופך להיות בלתי אפשרי להבחין בנפרד. ספירליזציה ודיספירליזציה קשורות לפעילות ה-DNA, מכיוון שהוא מתפקד רק במצב דה-פירליזציה. שחרור המידע, היווצרות RNA על DNA במצב ספירלי, כלומר בזמן מיטוזה, נעצרת. העובדה שכרומוזומים נמצאים בגרעין של תא שאינו מתחלק מוכחת גם על ידי קביעות כמות ה-DNA, מספר הכרומוזומים ושימור האינדיבידואליות שלהם מחלוקה לחלוקה.
הכנת תא למיטוזה. במהלך interphase מתרחשים מספר תהליכים המאפשרים מיטוזה. בואו נמנה את החשובים שבהם: 1) צנטריולים מוכפלים, 2) כרומוזומים מוכפלים, כלומר. כמות ה-DNA והחלבונים הכרומוזומליים, 3) מסונתזים חלבונים שמהם בנוי ציר האכרומטין, 4) נצברת אנרגיה בצורה של ATP, הנצרכת במהלך החלוקה, 5) מסתיימת צמיחת התאים. חשיבות עליונה בהכנת תא למיטוזה היא סינתזה של DNA ושכפול כרומוזומים. הכפלת הכרומוזומים קשורה בעיקר לסינתזה של DNA ולסינתזה סימולטנית של חלבוני כרומוזומים. תהליך ההכפלה אורך 6-10 שעות ואורך חלק אמצעיאינטרפאזות. שכפול הכרומוזומים מתנהל בצורה כזו שכל גדיל בודד ישן של DNA בונה לעצמו גדיל שני. תהליך זה מסודר בקפדנות, והחל ממספר נקודות, מתפשט לאורך כל הכרומוזום.
מיטוזה
מיטוזיס היא שיטה אוניברסלית לחלוקת תאים בצמחים ובעלי חיים, שמהותה העיקרית היא הפיזור המדויק של כרומוזומים משוכפלים בין שני תאי הבת שנוצרו. הכנת תא לחלוקה, כפי שאנו יכולים לראות, תופסת חלק משמעותי מהשלב הבין-פאזי, והמיטוזה מתחילה רק כאשר ההכנה בגרעין ובציטופלזמה הושלמה לחלוטין. כל התהליך מחולק לארבעה שלבים. במהלך הראשון שבהם - פרופסה - מתחלקים צנטריולים ומתחילים להתפצל לכיוונים מנוגדים. סביבם נוצרים חוטי אכרומטין מהציטופלזמה, אשר יחד עם צנטריולים יוצרים ציר אכרומטין. כאשר מסתיימת ההתפצלות של הצנטריולים, התא כולו קוטבי, שני הצנטריולים ממוקמים בקטבים מנוגדים, וניתן לקרוא למישור האמצעי קו המשווה. החוטים של ציר האכרומטין מתכנסים בצנטריולים ומופצים באופן נרחב בקו המשווה, בדומה לציר בצורתו. במקביל להיווצרות ציר בציטופלזמה, הגרעין מתחיל להתנפח, ובו מובחן בבירור כדור של חוטים מעובים - כרומוזומים. במהלך הפרופזה, הכרומוזומים מתגלגלים, מתקצרים ומתעבים. פרופאזה מסתיימת בפירוק המעטפת הגרעינית, ונמצא שהכרומוזומים שוכבים בציטופלזמה. בשלב זה, ניתן לראות שכל הכרומוזומים כבר כפולים. ואז מגיע השלב השני - מטאפאזה. כרומוזומים, המסודרים באקראי בהתחלה, מתחילים לנוע לעבר קו המשווה. כולם ממוקמים בדרך כלל באותו מישור במרחק שווה מהצנטריולים. בשלב זה חלק מחוטי הציר מחובר לכרומוזומים, בעוד שחלקם השני עדיין נמתח ברציפות מצנטרול אחד למשנהו - אלו הם החוטים התומכים. חוטים מושכים, או כרומוזומליים, מחוברים לצנטרומרים (התכווצויות ראשוניות של כרומוזומים), אך יש לזכור שגם הכרומוזומים וגם הצנטרומרים כבר כפולים. חוטי משיכת מהקטבים מחוברים לאותם כרומוזומים הקרובים יותר אליהם. יש הפסקה קצרה. זה חלק מרכזימיטוזה, ולאחריה מתחיל השלב השלישי - אנפאזה. במהלך אנפאזה, חוטי המשיכה של הציר מתחילים להתכווץ, מותחים את הכרומוזומים לקטבים שונים. במקרה זה, הכרומוזומים מתנהגים באופן פסיבי, הם מתכופפים כמו סיכת ראש, נעים קדימה על ידי צנטרומרים, שעבורם הם נמשכים על ידי חוט ציר. בתחילת האנפאזה, צמיגות הציטופלזמה יורדת, מה שתורם לתנועה מהירה של כרומוזומים. כתוצאה מכך, חוטי הציר מבטיחים את ההתרחקות המדויקת של הכרומוזומים (הכפלה אפילו ב-interphase) לקטבים שונים של התא. מיטוזה מסתיימת בשלב האחרון, טלופאז. כרומוזומים, המתקרבים לקטבים, שלובים זה בזה באופן הדוק. במקביל, מתחילה המתיחה (דיספיראליזציה) שלהם, ואי אפשר להבחין בין כרומוזומים בודדים. בהדרגה נוצרת המעטפת הגרעינית מהציטופלזמה, הגרעין מתנפח, הגרעין מופיע ומשחזר המבנה הקודם של העצמי הבין-פאזי.
חלוקת תאים מיטוטיים
מיטוזה(מיוונית. מיטוס - חוט), הנקרא גם קריוקינזיס, או חלוקת תאים עקיפה, הוא מנגנון אוניברסלי לחלוקת תאים. מיטוזיס עוקבת אחר תקופת G2 ומשלימה את מחזור התא.
זה נמשך 1-3 שעות ומבטיח פיזור אחיד של החומר הגנטי לתאי הבת. למיטוזה יש 4 שלבים עיקריים: פרופאזה, מטאפאזה, אנפאזה וטלופזה.
מיטוזה היא אחד התהליכים הבסיסיים של אונטוגניה. חלוקה מיטוטית מבטיחה צמיחה של איקריוטים רב-תאיים על ידי הגדלת אוכלוסיות תאי הרקמה.
כתוצאה מחלוקה מיטוטית של תאי מריסטם, מספר תאי רקמת הצמח עולה. פיצול ביצית מופרית וצמיחת רוב הרקמות בבעלי חיים מתרחשים גם באמצעות חלוקות מיטוטיות.
על בסיס תכונות מורפולוגיות, מיטוזה מחולקת לשלבים על תנאי: פרופאזה, פרומטפאזה, מטאפאזה, אנפאזה, טלופזה. התיאורים הראשונים של שלבי המיטוזה והקמת הרצף שלהם בוצעו בשנות ה-70-80 של המאה ה-19. בסוף שנות ה-70, ההיסטולוג הגרמני וולטר פלמינג טבע את המונח "מיטוזה" כדי להתייחס לתהליך של חלוקת תאים עקיפה.
משך המיטוזה הממוצע הוא 1-2 שעות. מיטוזה של תאי בעלי חיים, ככלל, נמשכת 30-60 דקות, וצמחים - 2-3 שעות. במשך 70 שנה, כ-10 14 חלוקות תאים מבוצעות בגוף האדם בסך הכל.
התיאורים הלא שלמים הראשונים הנוגעים להתנהגות ושינויים של גרעינים בתאים מתחלקים נמצאים בעבודותיהם של מדענים בתחילת שנות ה-70.
בעבודתו של הבוטנאי הרוסי רוסוב, משנת 1872, מתוארים ומתוארים בבירור לוחות מטאפאזה ואנפאזה, המורכבים מכרומוזומים בודדים.
שנה לאחר מכן, הזואולוג הגרמני G.A. שניידר אפילו בצורה ברורה ועקבית יותר, אבל, כמובן, לא לגמרי תיאר את החלוקה המיטוטית באמצעות הדוגמה של ריסוק ביצים של ה-Turbellaria Mesostomum פי הטבעת. בעבודתו, בעצם, מתוארים ומאוירים השלבים העיקריים של המיטוזה ברצף הנכון: פרופאזה, מטאפאזה, אנפאזה (מוקדמת ומאוחרת). בשנת 1874, הבוטנאי של מוסקבה I.D. צ'יסטיאקוב צפה גם בשלבים נפרדים של חלוקת תאים בנבגים של טחב מועדון וזנב סוס. למרות ההצלחות הראשונות, לא רוסוב, לא שניידר ולא צ'יסטיאקוב הצליחו לתת תיאור ברור ועקבי של החלוקה המיטוטית.
בשנת 1875 יצאו יצירות המכילות יותר מ תיאורים מפורטיםמיטוזות. O. Byuchli נתן תיאור של דפוסים ציטולוגיים בריסוק הביצים של תולעים עגולות ורכיכות ובתאים spermatogenic של חרקים.
E. Strasburger חקר חלוקה מיטוטית בתאי האצה הירוקה spirogyra, בתאי האם של אבקת הבצל, ובתאי נבגי האם של אזוב המועדון. בהתייחס לעבודתו של O. Buechli ועל סמך מחקרו שלו, E. Strasburger הפנה את תשומת הלב לאחדות תהליכי חלוקת התאים בתאי צמחים ובעלי חיים.
עד סוף 1878 - תחילת 1879 הופיעו יצירות מפורטות של שלייכר ושל וו. פלמינג. בעבודתו בשנת 1879, הציע שלייכר את המונח "קריוקינזיס" כדי להתייחס לתהליכים המורכבים של חלוקת תאים, מה שמרמז על התנועה חלקי מרכיביםגרעינים. וולטר פלמינג היה הראשון שהציג את המונח "מיטוזה" המתייחס לחלוקת תאים עקיפה, שהפכה מאוחר יותר למקובלת. פלמינג הוא גם הבעלים של הניסוח הסופי של ההגדרה של מיטוזה כתהליך מחזורי, ששיאו בחלוקת כרומוזומים בין תאי בת.
בשנת 1880 O.V. ברנצקי קבע את המבנה הסליל של הכרומוזומים. במהלך מחקר נוסף, פותחו רעיונות לגבי הספירליזציה והדיספירליזציה של כרומוזומים במהלך המחזור המיטוטי.
בתחילת שנות ה-1900, כרומוזומים זוהו כנשאים של מידע תורשתי, מה שמאוחר יותר סיפק הסבר תפקיד ביולוגימיטוזה, המורכבת מיצירת תאי בת זהים מבחינה גנטית.
בשנות ה-70 החל הפענוח והמחקר המפורט של מווסת החלוקה המיטוטית, הודות לסדרת ניסויים על היתוך תאים הממוקמים על שלבים שוניםמחזור התא. באותם ניסויים, כאשר תא בשלב M שולב עם תא בכל אחד משלבי האינטרפאזה (G1, S או G2), עברו התאים הבין-פאזיים למצב מיטוטי (התחיל עיבוי כרומוזומים והמעטפת הגרעינית התפרקה) .
כתוצאה מכך, הגיעו למסקנה שיש גורם (או גורמים) בתא הציטופלזמי הממריץ מיטוזה, או, במילים אחרות, הגורם המגרה M (MSF, מה-M-phase-promoting factor באנגלית, MPF) .
לראשונה התגלה "הגורם המעורר מיטוזיס" בביציות לא מופרות בשלות של הצפרדע בעלת הטפרים, שנמצאות ב-M-phase של מחזור התא. הציטופלזמה של ביצית כזו, שהוזרקה לביצית, הובילה למעבר מוקדם ל-M-phase ולהתחלת הבשלה של הביצית (במקור, הקיצור MPF עמד על Maturation Promoting Factor, שמתורגם כ"התבגרות גורם מקדם"). במהלך ניסויים נוספים, הובאו המשמעות האוניברסלית, ובמקביל, רמה גבוהה של שמרנות של "הגורם הממריץ מיטוזיס": תמציות שהוכנו מתאי מיטוטיים של אורגניזמים מגוונים מאוד, כשהם מוכנסים לתוך ביציות צפרדע עם טפרים, הועברו. אותם לשלב M.
מחקרים שלאחר מכן גילו כי הגורם הממריץ מיטוזה הוא קומפלקס הטרודימרי המורכב מחלבון ציקלין וחלבון קינאז תלוי ציקלין. ציקלין הוא חלבון רגולטורי ונמצא בכל האיקריוטים. ריכוזו עולה מעת לעת במהלך מחזור התא, ומגיע למקסימום במטאפאזה של מיטוזה. עם תחילת האנפאזה, נצפית ירידה חדה בריכוז הציקלין, עקב ביקוע שלו בעזרת קומפלקסים פרוטאוליטיים מורכבים של חלבון - פרוטאוזומים. חלבון קינאז תלוי ציקלין הוא אנזים (פוספורילאז) שמשנה חלבונים על ידי העברת קבוצת פוספטים מ-ATP לחומצות האמינו סרין ותריונין. כך, עם ביסוס התפקיד והמבנה של הרגולטור הראשי של החלוקה המיטוטית, החלו מחקרים על מנגנוני הוויסות העדינים של המיטוזה, הנמשכים עד היום.
הפיתוח של טיפולוגיה וסיווג מאוחדים של מיטוזות מסובך על ידי מגוון שלם של תכונות אשר, בשילובים שונים, יוצרות מגוון והטרוגניות של דפוסים של חלוקה מיטוטית. יחד עם זאת, אפשרויות סיווג נפרדות שפותחו עבור טקסות מסוימות אינן מקובלות על אחרות, מכיוון שהן אינן לוקחות בחשבון את הספציפיות של המיטוזה שלהן. לדוגמה, כמה גרסאות של סיווג מיטוזות, האופייניות לאורגניזמים של בעלי חיים או צמחיים, מתבררות כבלתי מקובלות עבור אצות.
אחד מ תכונות עיקריותבבסיס הטיפולוגיות והסיווגים השונים של החלוקה המיטוטית עומדת ההתנהגות של המעטפת הגרעינית. אם היווצרות הציר והחלוקה המיטוטית עצמה ממשיכה בתוך הגרעין מבלי להרוס את הממברנה הגרעינית, אז סוג זה של מיטוזה נקרא סגור. מיטוזה עם קריסת המעטפת הגרעינית, בהתאמה, נקראת פתוחה, ומיטוזה עם קריסת הממברנה רק בקטבים של הציר, עם היווצרות "חלונות קוטביים" - סגורים למחצה.
תכונה אופיינית נוספת היא סוג הסימטריה של הציר המיטוטי. בפלורומיטוזיס, ציר החלוקה הוא סימטרי או א-סימטרי דו-צדדית ובדרך כלל מורכב משני סמי-צירים הממוקמים במטאפאזה-אנפאזה בזווית זה לזה. קטגוריית האורתומיטוזות מאופיינת בסימטריה דו-קוטבית של ציר הביקוע, ובמטאפזה יש לרוב לוח משווני שניתן להבחין בו.
בתוך הסימנים המצוינים, הרב ביותר הוא אורתומיטוזיס פתוח טיפוסי, שעל הדוגמה שלו נדונים להלן העקרונות ושלבי החלוקה המיטוטית. הסוג הזהמיטוזה אופיינית לבעלי חיים, לצמחים גבוהים ולכמה פרוטוזואה.
פרופאזה מתחילה בהתעבות של כרומוזומים, הנראים תחת מיקרוסקופ אור כמבנים חוטיים. כל כרומוזום מורכב משתי כרומטידות אחיות מקבילות המחוברות בצנטרומר. הגרעין והמעטפת הגרעינית נעלמים עד סוף השלב (האחרונה מתפרקת לשלפוחיות ממברנות הדומות ליסודות ה-EPS, וקומפלקס הנקבוביות והלמינה מתפרקים ליחידות משנה). קריופלזמה מתערבבת עם ציטופלזמה.
צנטרולים נודדים לקטבים מנוגדים של התא ומולידים את החוטים של הציר המיטוטי (אכרומטין). באזור הצנטרומר נוצרים קומפלקסים מיוחדים של חלבון - קינטוכורים, אליהם מחוברות כמה מיקרוטובוליות ציר (kinetochore microtubules); הוכח כי קינטוכורים עצמם מסוגלים לגרום להרכבת מיקרוטובוליות ולכן יכולים לשמש כמרכזי ארגון מיקרוטובוליים. שאר מיקרו-צינוריות הציר נקראות מיקרו-צינוריות קוטביות, שכן הן משתרעות מקוטב אחד של התא למשנהו; המיקרוטובולים השוכבים מחוץ לציר, מתפצלים באופן רדיאלי ממרכזי התא אל הפלזמה, קיבלו את השם אסטרלי או מיקרוטובוליות (חוטים) של זוהר.
מטאפאזה תואמת רמה מקסימליתהתעבות של כרומוזומים המסתדרים באזור המשווני של הציר המיטוטי, ויוצרים תמונה של הלוח המשווני (מטאפזה) (מבט צד) או כוכב האב (מבט מהקטבים). כרומוזומים נעים למישור המשווני ומוחזקים בו בשל המתח המאוזן של המיקרוטובולים הקינטוכורים. בסוף שלב זה, כרומטידות אחיות מופרדות בפער, אך נשמרות באזור הצנטרומר.
אנפאזה מתחילה בפיצול סינכרוני של כל הכרומוזומים לכרומטידות אחיות (באזור הצנטרומר) ובתנועה של כרומוזומי הבת לקטבים מנוגדים של התא, המתרחשת לאורך המיקרוטובוליות בציר במהירות של 0.2-0.5 מיקרומטר/דקה. האות להופעת אנפאזה כולל עלייה חדה (בסדר גודל) בריכוז קטיוני הסידן בהיאלופלזמה, המופרשת על ידי שלפוחיות ממברנות היוצרות צבירים בקוטבי הציר. מנגנון תנועת הכרומוזומים באנפאזה לא הובהר במלואו, אולם, הוכח כי בנוסף לאקטין, חלבונים כגון מיוזין ודיניין, כמו גם מספר חלבונים מווסתים, נמצאים באזור הציר. על פי כמה תצפיות, זה נובע מקיצור (פירוק) של מיקרוטובולים המחוברים לקינטוכורים. אנפאזה מאופיינת בהתארכות של הציר המיטוטי עקב התבדלות מסוימת של קטבי התא. זה מסתיים בהצטברות של שתי קבוצות זהות של כרומוזומים בקטבים של התא, היוצרים תמונות של כוכבים (שלב כוכבי הבת). בתום האנפאזה, עקב התכווצות מיקרופילמנטים של אקטין, המתרכזים סביב היקף התא (טבעת מתכווצת), מתחילה להיווצר התכווצות תא, אשר מעמיקה תוביל בשלב הבא לציטוטומיה.
טלופאז הוא שלב סופימיטוזה, שבמהלכה נבנים מחדש גרעיני תאי הבת והושלמה חלוקתם. מסביב לכרומוזומים המעובים של תאי הבת משלפוחיות ממברנות (לפי מקורות אחרים, מ-EPS), משוחזרת הקריולמה, שאליה קשורה הלמינה המתעוררת, חוזרים ומופיעים נוקלאולי, הנוצרים מקטעים של הכרומוזומים המקבילים. גרעיני התא גדלים בהדרגה, והכרומוזומים מתדרדרים בהדרגה ונעלמים, ומוחלפים בתבנית הכרומטין של גרעין הבין-פאזי. במקביל, התכווצות התא מעמיקה, והתאים נשארים מחוברים לזמן מה על ידי גשר ציטופלזמי מצטמצם המכיל צרור של מיקרוטובולים (גוף אמצעי). קשירה נוספת של הציטופלזמה מסתיימת ביצירת שני תאי בת. בטלופאזה מתרחשת התפלגות האברונים בין תאי הבת; האחידות של תהליך זה מקלה על העובדה שחלק מהאברונים הם רבים למדי (למשל, מיטוכונדריה), בעוד שאחרים (כמו EPS ו-Golgi קומפלקס) מתפרקים לשברים קטנים ושלפוחיות במהלך מיטוזה.
מיטוזות לא טיפוסיות מתרחשות כאשר המנגנון המיטוטי ניזוק ומאופיינות בפיזור לא אחיד של חומר גנטי בין תאים - אנופלואידיה (מיוונית an - not, eu - correct, ploon - add); במקרים רבים, ציטוטומיה נעדרת, וכתוצאה מכך נוצרים תאים ענקיים. מיטוזות לא טיפוסיות אופייניות לגידולים ממאירים ורקמות מוקרנות. ככל שתדירותם גבוהה יותר וככל שמידת האנופלואידיה גדולה יותר, כך הגידול ממאיר יותר. הפרה של חלוקת תאים מיטוטית נורמלית יכולה להיגרם על ידי חריגות כרומוזומים, אשר נקראות סטיות כרומוזומליות (מלטינית Aberratio - סטייה). גרסאות של סטיות כרומוזומליות הן הידבקות של כרומוזומים, קרע שלהם לרסיסים, אובדן אתר, החלפת שברים, הכפלה סעיפים בודדיםכרומוזומים וכו'. סטיות כרומוזומליות יכולות להתרחש באופן ספונטני, אך לעתים קרובות יותר הן מתפתחות כתוצאה מפעולת מוטגנים וקרינה מייננת על תאים.
קריוטיפינג - מחקר אבחוני להערכת הקריוטיפ (מערכת הכרומוזומים) מתבצע על ידי בדיקת הכרומוזומים בצלחת המטאפאזה. לצורך קריוטיפינג מתקבלת תרבית תאים שאליה מכניסים קולכיצין, חומר החוסם את היווצרות הציר המיטוטי. כרומוזומים מופקים מתאי כאלה, אשר נצבעים ומזוהים עוד יותר. הקריוטיפ האנושי הנורמלי מיוצג על ידי 46 כרומוזומים - 22 זוגות של אוטוזומים ושני כרומוזומי מין (XY בגברים ו-XX בנשים). קריוטיפינג מאפשר לאבחן מספר מחלות הקשורות להפרעות כרומוזומליות, בפרט, תסמונת דאון (טריזומיה של הכרומוזום ה-21), אדוארדס (טריזומיה של הכרומוזום ה-18), פטאו (טריזומיה של הכרומוזום ה-13), וכן מספר תסמונות הקשורות לאנומליות כרומוזומי מין - תסמונת קלינפלטר (גנוטיפ - XXY), טרנר (גנוטיפ - XO) ואחרים.
ההנחה היא שהתהליך המיטוטי המורכב של אורגניזמים גבוהים יותר התפתח בהדרגה ממנגנוני החלוקה הפרוקריוטית. הנחה זו נתמכת על ידי העובדה כי פרוקריוטים הופיעו לפני כמיליארד שנים. לפני הראשוןאיקריוטים. בנוסף, חלבונים דומים מעורבים במיטוזה איקריוטית ובביקוע בינארי פרוקריוטי.
ניתן לאתר שלבי ביניים אפשריים בין ביקוע בינארי למיטוזה באאוקריוטים חד-תאיים, שבהם הקרום הגרעיני אינו נהרס במהלך החלוקה. ברוב האיקריוטים האחרים, כולל צמחים ובעלי חיים, ציר הביקוע נוצר מחוץ לגרעין, והממברנה הגרעינית נהרסת במהלך המיטוזה. למרות שהמיטוזה באאוקריוטים חד-תאיים עדיין לא מובנת היטב, ניתן להניח שהיא מקורה בביקוע בינארי ולבסוף הגיעה לרמת המורכבות הקיימת באורגניזמים רב-תאיים.
באאוקריוטים פרוטוזואים רבים, המיטוזה נשארה גם תהליך הקשור לממברנה, אך כעת היא כבר לא פלזמה, אלא גרעינית.
מנגנוני הרגולציה העיקריים של מיטוזה הם תהליכי הזרחון והפרוטאוליזה.
תגובות זרחון ודפוספורילציה הפיכות מאפשרות אירועים מיטוטיים הפיכים כגון הרכבה/פירוק ציר או פירוק/תיקון מעטפת גרעינית. פרוטאוליזה עומדת בבסיס האירועים הבלתי הפיכים של מיטוזה, כגון הפרדת כרומטידות אחיות באנפאזה או פירוק של ציקלינים מיטוטיים ל שלבים מאוחריםמיטוזה.
החלוקה של כל התאים האיקריוטים קשורה להיווצרות של מנגנון מיוחד לחלוקת תאים.
תפקיד פעיל בחלוקת תאים מיטוטיים מוקצה לעתים קרובות למבנים ציטו-שלד. הציר המיטוטי הדו-קוטבי, המורכב ממיקרו-צינוריות וחלבונים קשורים, הוא אוניברסלי עבור תאי בעלי חיים וצמחים כאחד. ציר החלוקה מספק חלוקה זהה לחלוטין של כרומוזומים בין קטבי החלוקה, שבאזורם נוצרים גרעיני תאי הבת בטלופאז.
תהליך המיטוזה מבטיח חלוקה אחידה למהדרין של כרומוזומים בין שני גרעיני בת, כך שבאורגניזם רב-תאי לכל התאים יש בדיוק את אותן קבוצות (במספר ובאופי) של כרומוזומים.
כרומוזומים מכילים מידע גנטי המקודד ב-DNA, ולכן תהליך מיטוטי קבוע ומסודר מבטיח גם העברה מלאה של כל המידע לכל אחד מגרעיני הבת; כתוצאה מכך, לכל תא יש את כל המידע הגנטי הדרוש לפיתוח כל המאפיינים של האורגניזם. בהקשר זה, מתברר מדוע תא אחד שנלקח מצמח בוגר מובחן לחלוטין יכול תנאים מתאימיםלהתפתח לצמח שלם. תיארנו מיטוזה ב תא דיפלואידי, אך תהליך זה מתרחש באופן דומה בתאים הפלואידים, למשל, בתאים של הצמחים מדור הגמטופיטים.